

A requirement for cyclin-dependent kinase 6 in thymocyte development and tumorigenesis
- PMID: 19155308
- PMCID: PMC2636510
- DOI: 10.1158/0008-5472.CAN-08-2473
A requirement for cyclin-dependent kinase 6 in thymocyte development and tumorigenesis
Abstract
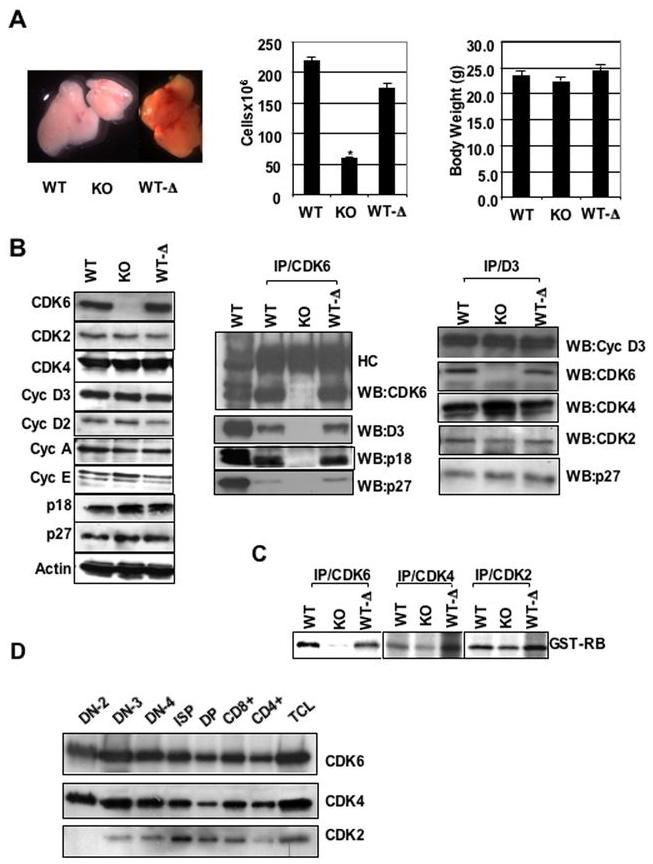
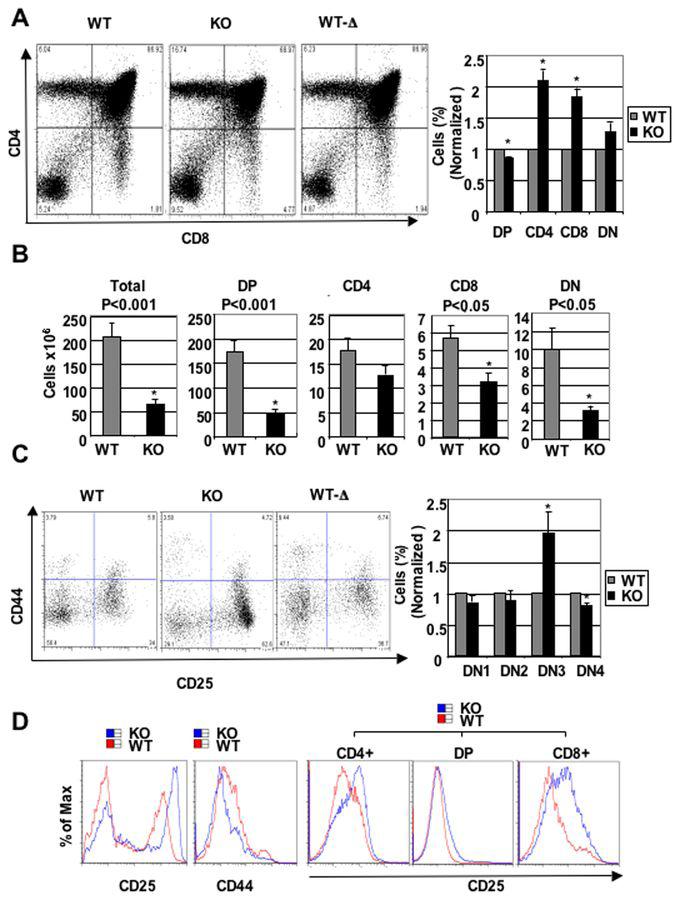
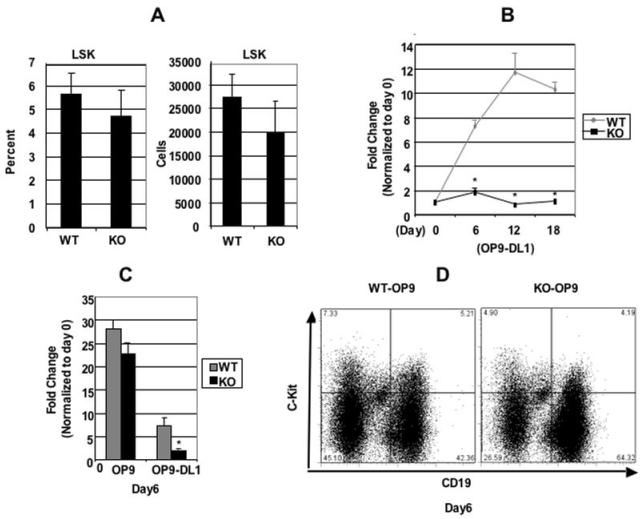
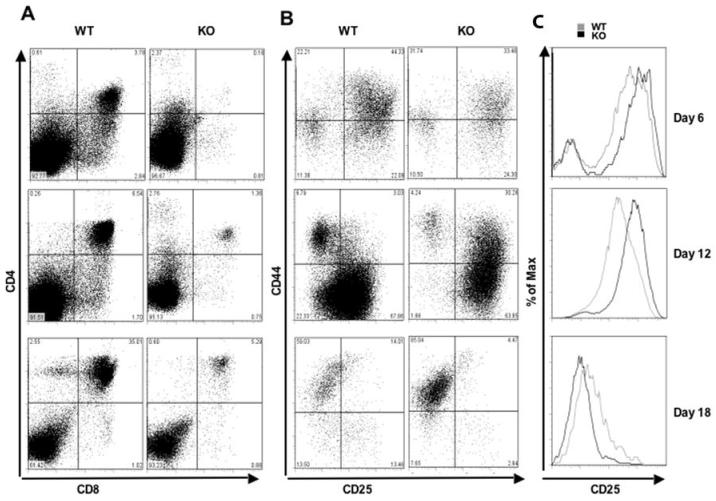
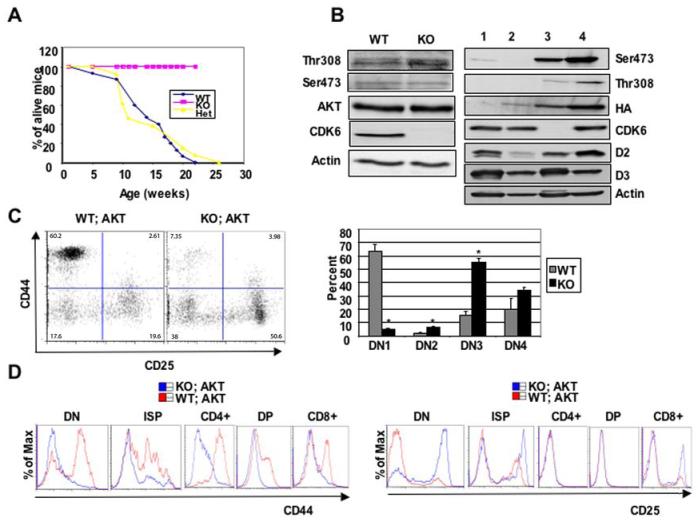
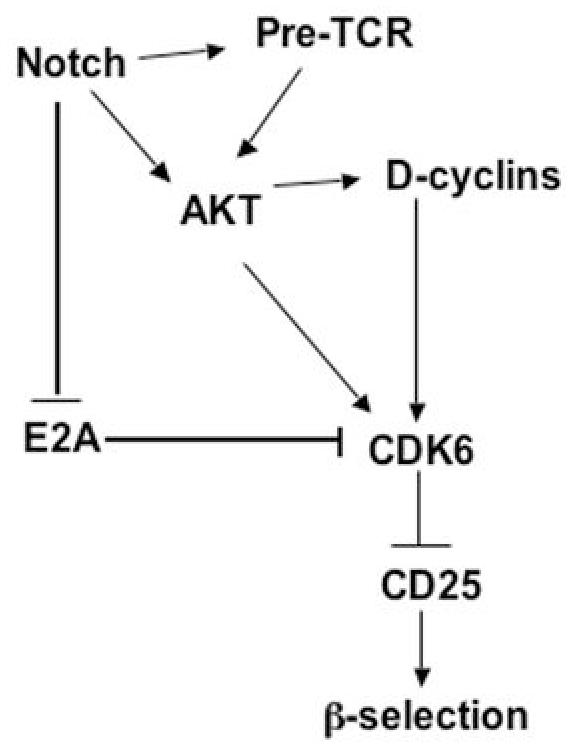
Cyclin-dependent kinase 6 (CDK6) promotes cell cycle progression and is overexpressed in human lymphoid malignancies. To determine the role of CDK6 in development and tumorigenesis, we generated and analyzed knockout mice. Cdk6-deficient mice show pronounced thymic atrophy due to reduced proliferative fractions and concomitant transitional blocks in the double-negative stages. Using the OP9-DL1 system to deliver temporally controlled Notch receptor-dependent signaling, we show that CDK6 is required for Notch-dependent survival, proliferation, and differentiation. Furthermore, CDK6-deficient mice were resistant to lymphomagenesis induced by active Akt, a downstream target of Notch signaling. These results show a critical requirement for CDK6 in Notch/Akt-dependent T-cell development and tumorigenesis and strongly support CDK6 as a specific therapeutic target in human lymphoid malignancies.
Figures
References
-
- Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fate control and signal integration in development. Science. 1999;284:770–6. - PubMed
-
- Radtke F, Wilson A, Stark G, et al. Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity. 1999;10:547–58. - PubMed
-
- Schmitt TM, Zuniga-Pflucker JC. Induction of T cell development from hematopoietic progenitor cells by delta-like-1 in vitro. Immunity. 2002;17:749–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
