

OX40-enhanced tumor rejection and effector T cell differentiation decreases with age
- PMID: 19155495
- PMCID: PMC6760259
- DOI: 10.4049/jimmunol.182.3.1481
OX40-enhanced tumor rejection and effector T cell differentiation decreases with age
Abstract
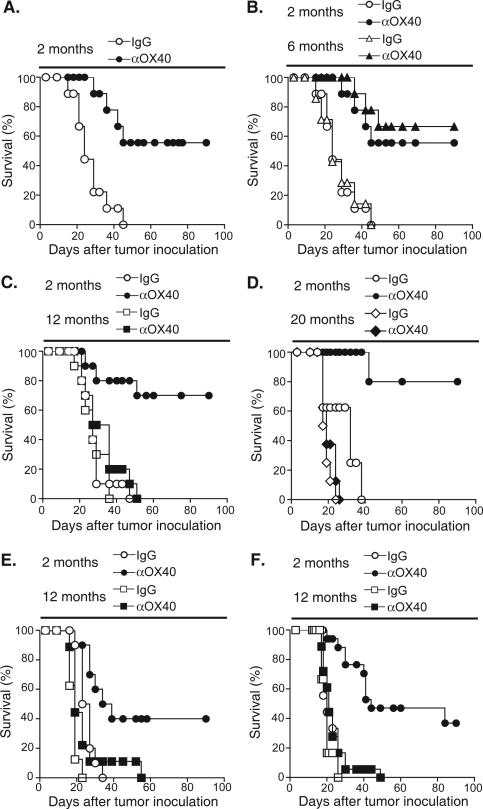
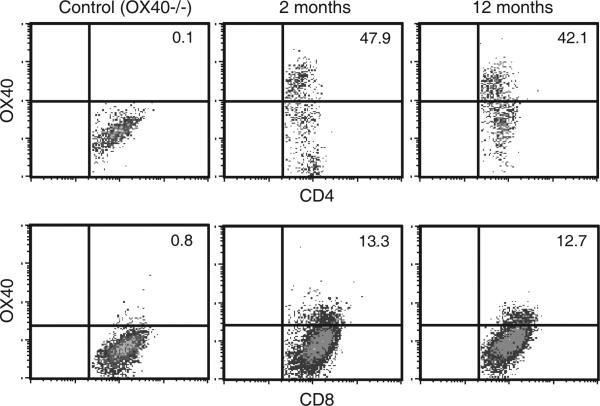
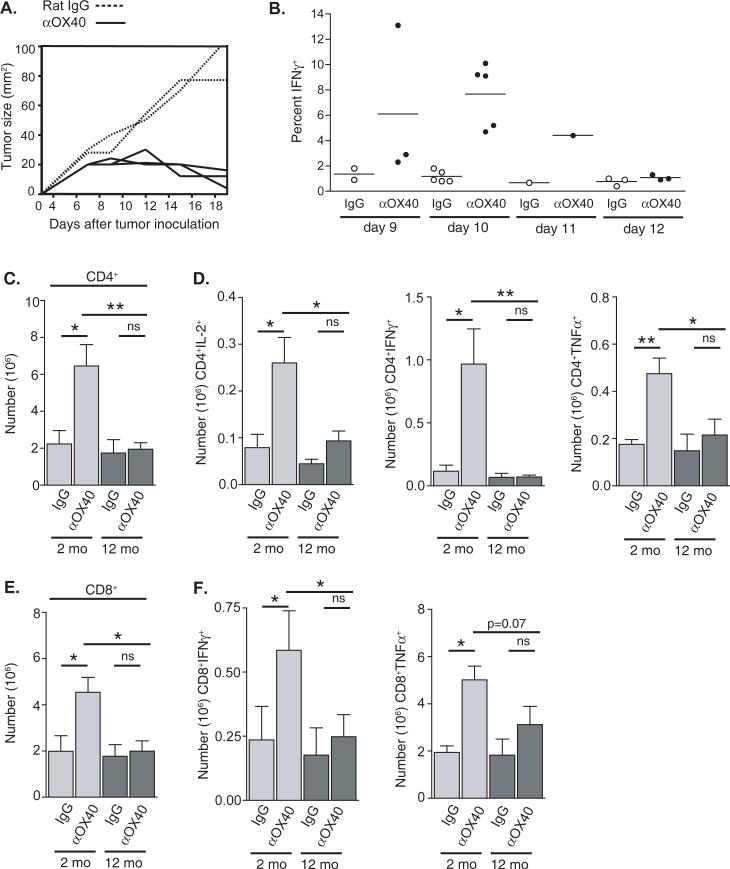
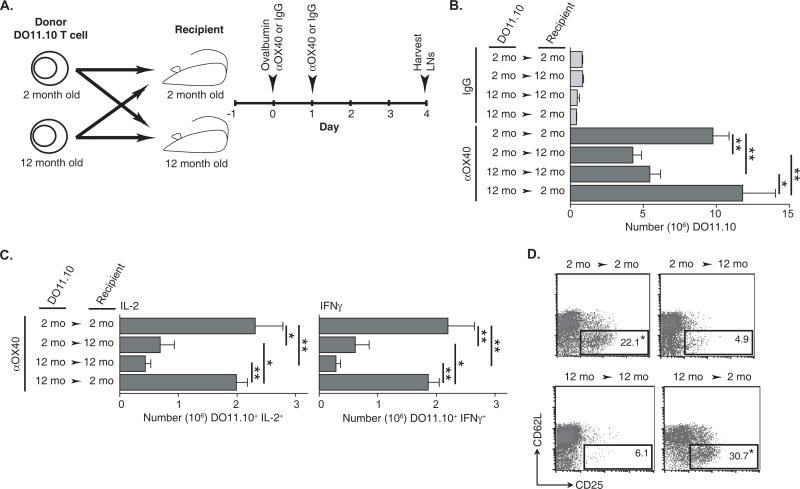
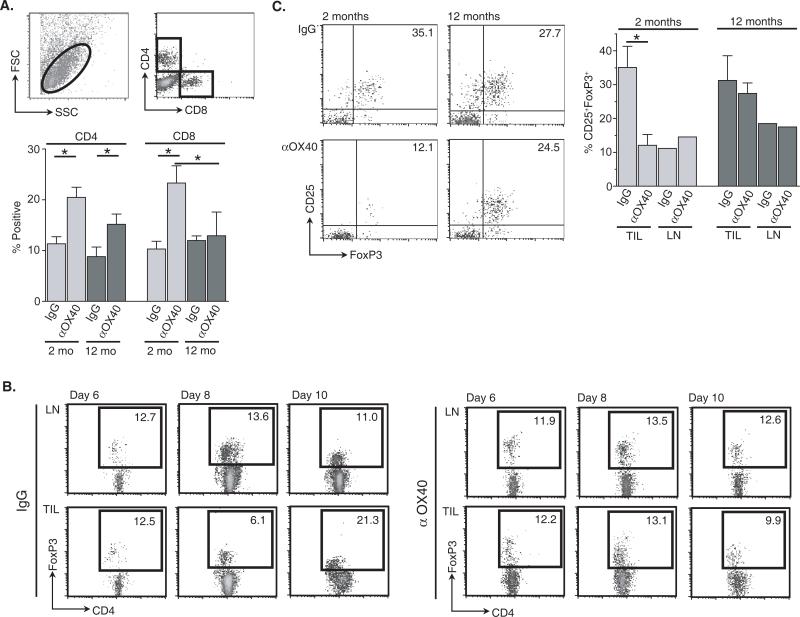
OX40 agonists have potent immunotherapeutic effects against a variety of murine tumors, yet it is unclear the role that age-related immune senescence plays on their efficacy. We found that middle-aged and elderly tumor-bearing mice (12 and 20 mo old, respectively) treated with anti-OX40 were less responsive compared with young mice 6 mo or less of age. Decreased tumor-free survival was observed in both male and female mice, and was not due to changes in the surface expression of OX40 on T cells in older animals. Enumeration of cytokine-producing effector T cells in tumor-bearing mice revealed a significant decline in these cells in the older mice treated with anti-OX40 compared with their younger counterparts. The decrease of this critical T cell population in middle-aged mice was not a result of inherent T cell deficiencies, but was revealed to be T cell extrinsic. Finally, combining IL-12, an innate cytokine, with anti-OX40 boosted levels of differentiated effector T cells in the older anti-OX40-treated mice and partially restored the defective antitumor responses in the middle-aged mice. Our data show that the anti-OX40-enhancement of tumor immunity and effector T cell numbers is decreased in middle-aged mice and was partially reversed by coadministration of the proinflammatory cytokine IL-12.
Figures


References
-
- Ershler WB. The influence of an aging immune system on cancer incidence and progression. J Gerontol. 1993;48:B3–7. - PubMed
-
- Yung RL. Changes in immune function with age. Rheum Dis Clin North Am. 2000;26:455–473. - PubMed
-
- Linton PJ, Haynes L, Tsui L, Zhang X, Swain S. From naive to effector--alterations with aging. Immunol Rev. 1997;160:9–18. - PubMed
-
- Effros RB, Walford RL. The immune response of aged mice to influenza: diminished T-cell proliferation, interleukin 2 production and cytotoxicity. Cell Immunol. 1983;81:298–305. - PubMed
-
- Melero I, Shuford WW, Newby SA, Aruffo A, Ledbetter JA, Hellstrom KE, Mittler RS, Chen L. Monoclonal antibodies against the 4-1BB T-cell activation molecule eradicate established tumors. Nat Med. 1997;3:682–685. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
