

The Cdk5/p35 kinases modulate leptin-induced STAT3 signaling
- PMID: 19156541
- PMCID: PMC2745521
- DOI: 10.1007/s12031-008-9174-3
The Cdk5/p35 kinases modulate leptin-induced STAT3 signaling
Abstract
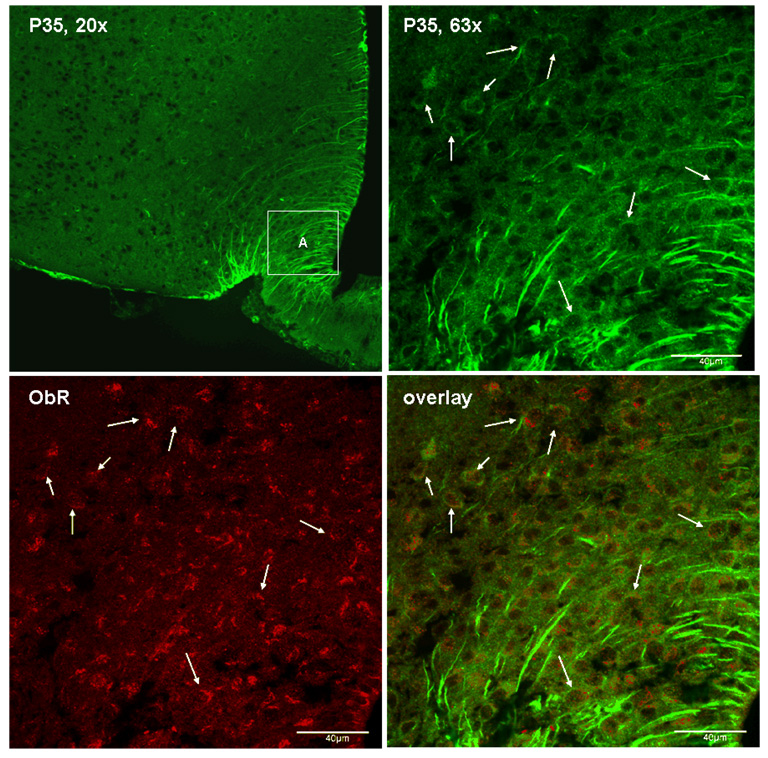
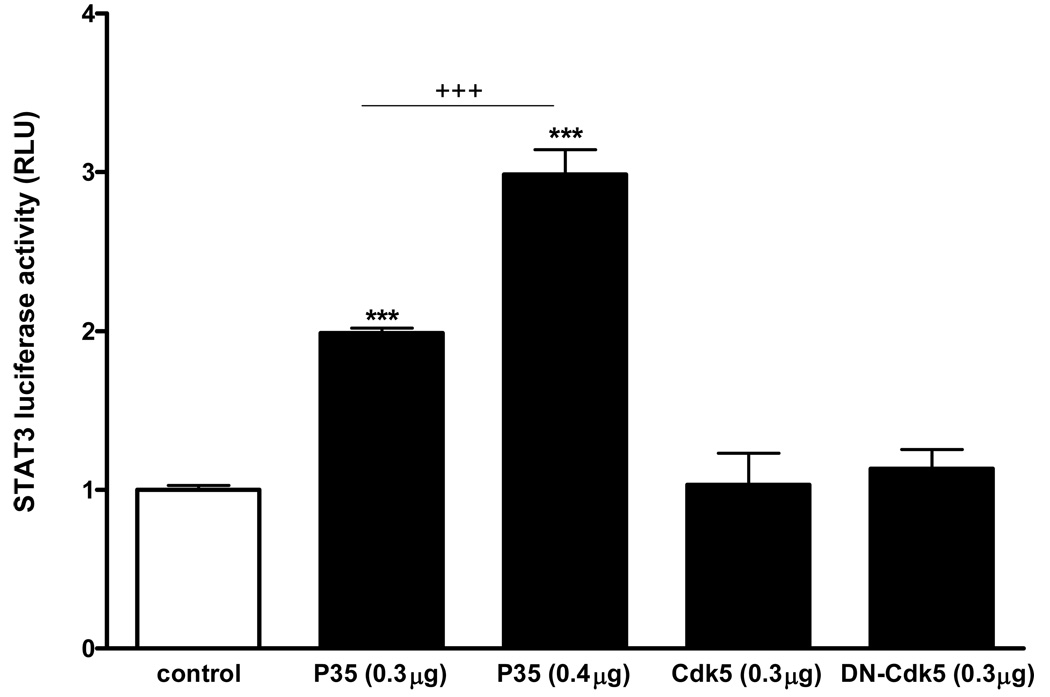
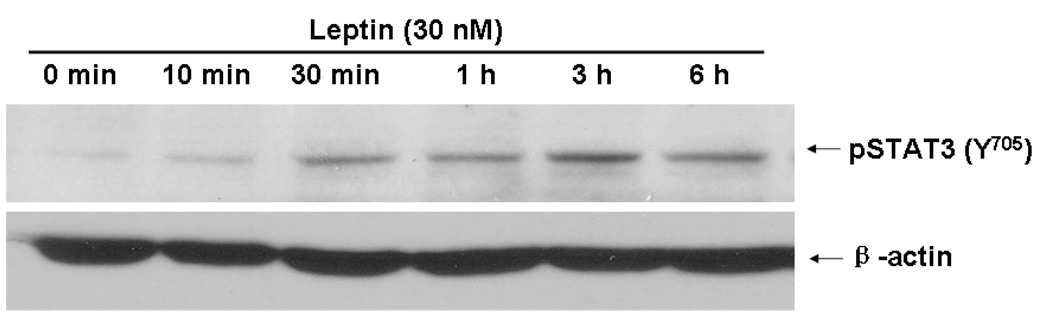
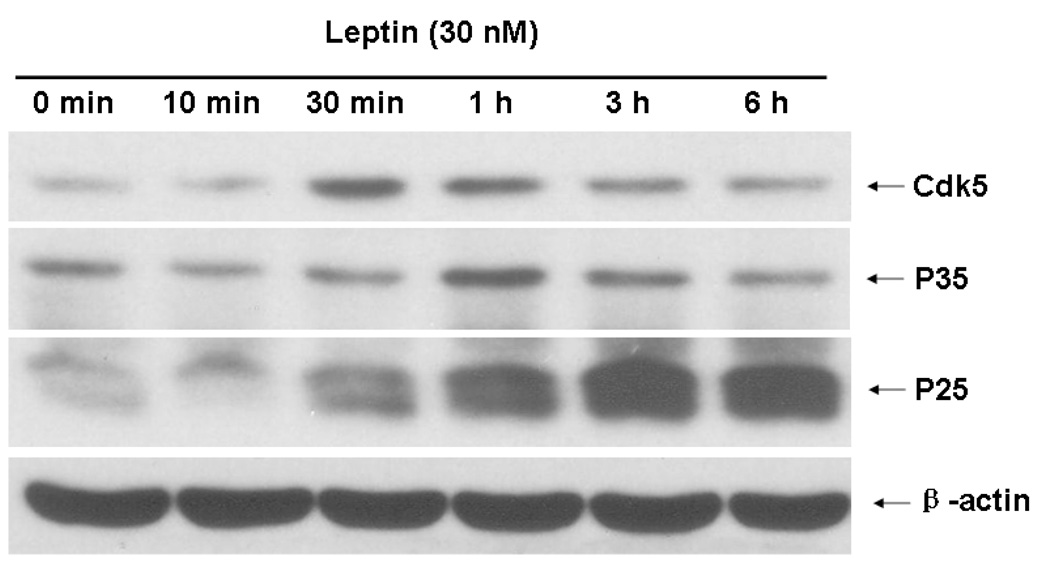
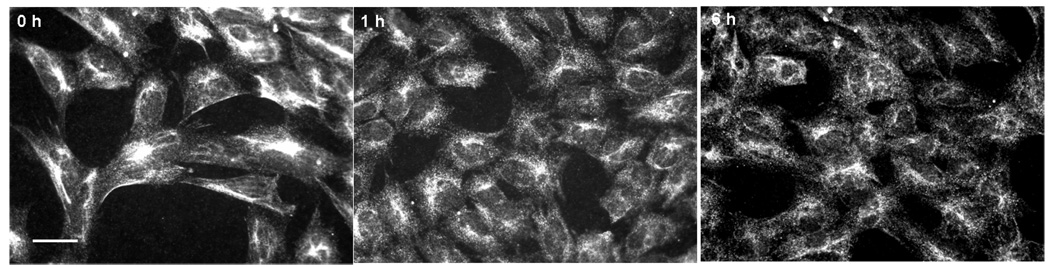
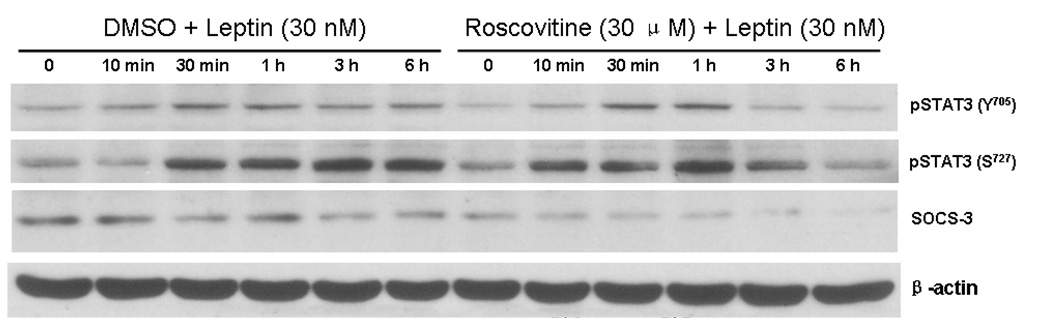
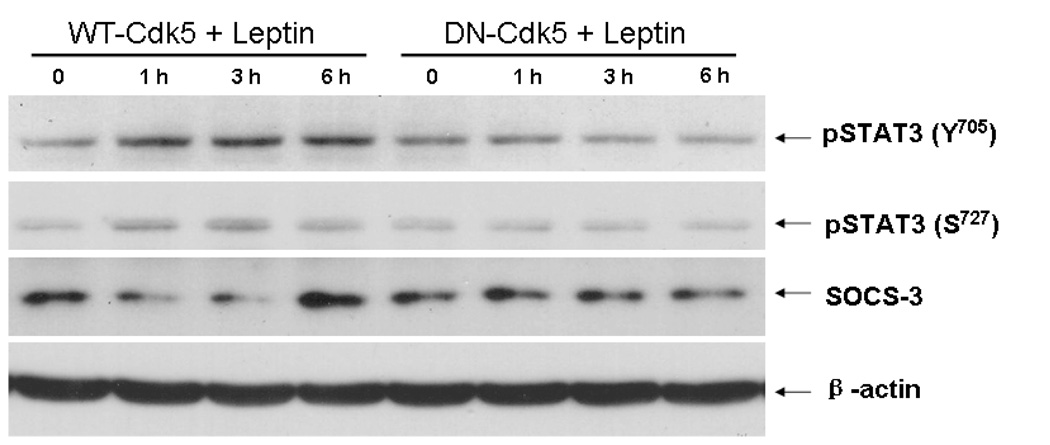
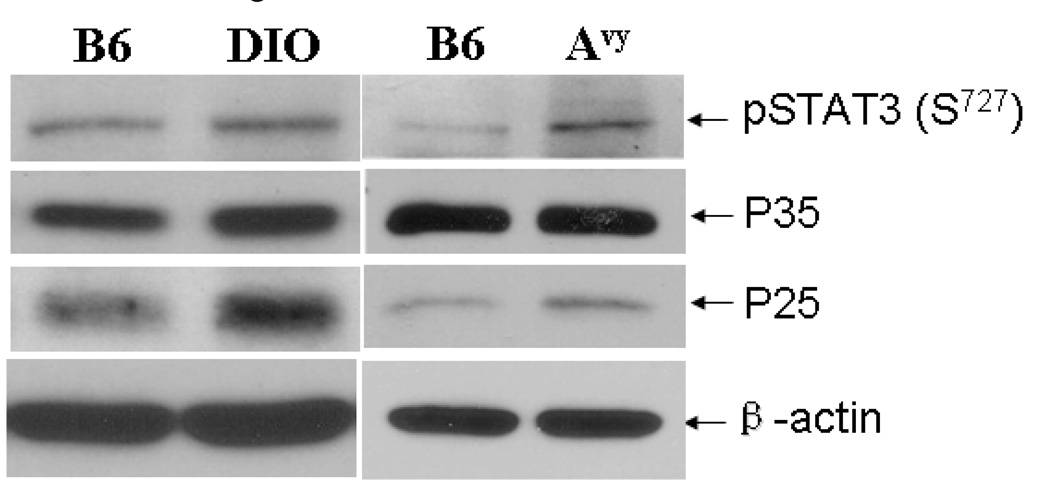
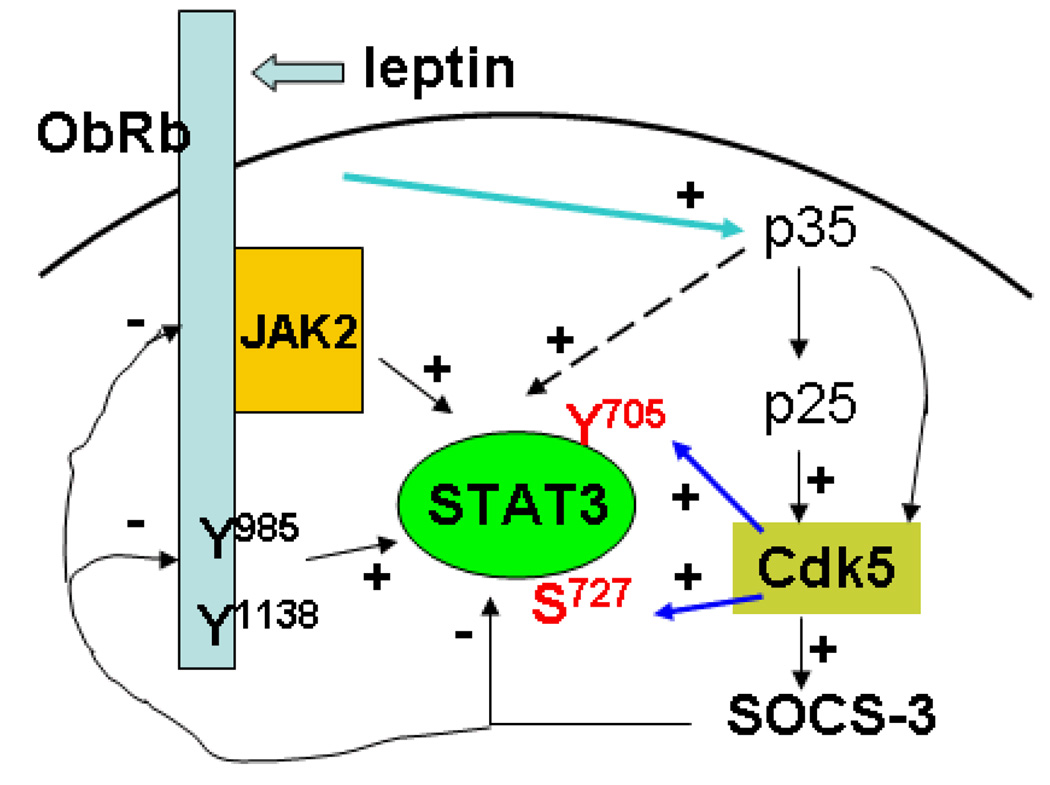
Cyclin-dependent kinase (Cdk) 5 is ubiquitously expressed in the brain and plays an essential role in central nervous system development and synaptic plasticity. The p35 kinase is a neuronal specific activator of Cdk5. Here, we show for the first time that Cdk5 activation modulates leptin signaling. P35 and its metabolite p25 were colocalized with the leptin receptor ObR in selective neurons in the hypothalamus. Overexpression of p35 alone was sufficient to induce the transcriptional activation of signal transducer and activator of transcription 3 (STAT3) in a cellular model. In retinoic acid-differentiated SH-SY5Y neuronal cells where ObRb was induced, leptin increased the expression of Cdk5, p35, and p25 kinases. The time course of induction coincided with that of phosphorylated (p)-STAT3. When Cdk5 activity was inhibited, either by roscovitine or overexpression of dominant negative Cdk5, there was a reduction of pSTAT3 activation. The results show that the activation of Cdk5 by p35 sustained leptin-induced pSTAT3 at 3-6 h. Thus, p35 is a novel modulator of leptin-induced STAT3 signaling.
Figures


References
-
- Banks WA, Kastin AJ, Huang W, Jaspan JB, Maness LM. Leptin enters the brain by a saturable system independent of insulin. Peptides. 1996;17:305–311. - PubMed
-
- Baumann K, Mandelkow EM, Biernat J, Piwnica-Worms H, Mandelkow E. Abnormal Alzheimer-like phosphorylation of tau-protein by cyclin-dependent kinases cdk2 and cdk5. FEBS Lett. 1993;336:417–424. - PubMed
-
- Bjorbak C, Lavery HJ, Bates SH, Olson RK, Davis SM, Flier JS, Myers MG., Jr SOCS3 mediates feedback inhibition of the leptin receptor via Tyr985. J Biol Chem. 2000;275:40649–40657. - PubMed
-
- Cheung ZH, Fu AK, Ip NY. Synaptic roles of Cdk5: implications in higher cognitive functions and neurodegenerative diseases. Neuron. 2006;50:13–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
