

Structure-function relationship of the chloroplastic glutaredoxin S12 with an atypical WCSYS active site
- PMID: 19158074
- PMCID: PMC2666582
- DOI: 10.1074/jbc.M807998200
Structure-function relationship of the chloroplastic glutaredoxin S12 with an atypical WCSYS active site
Abstract
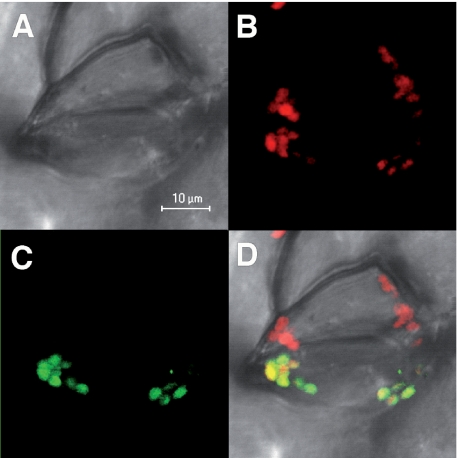
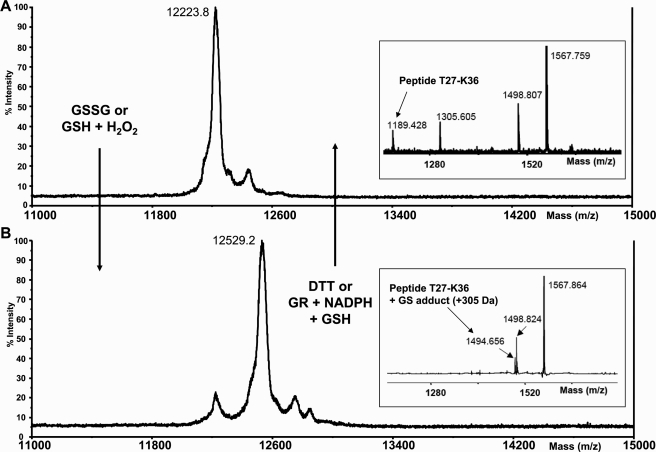
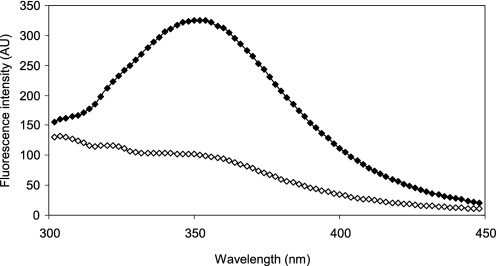
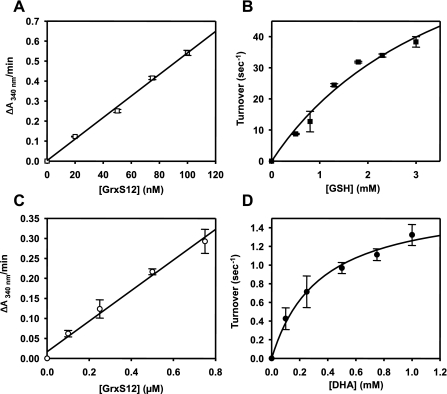
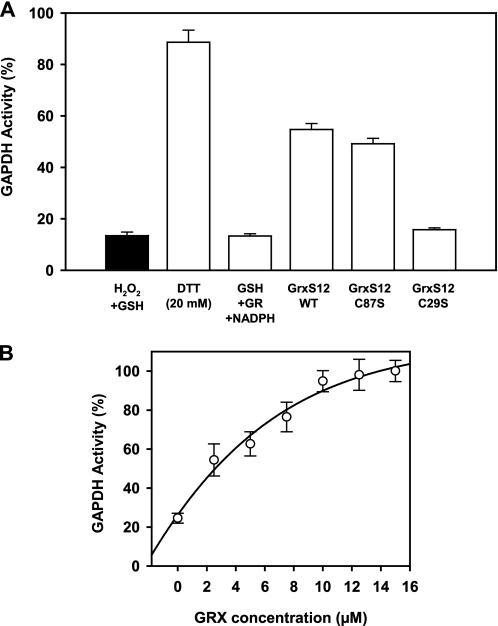
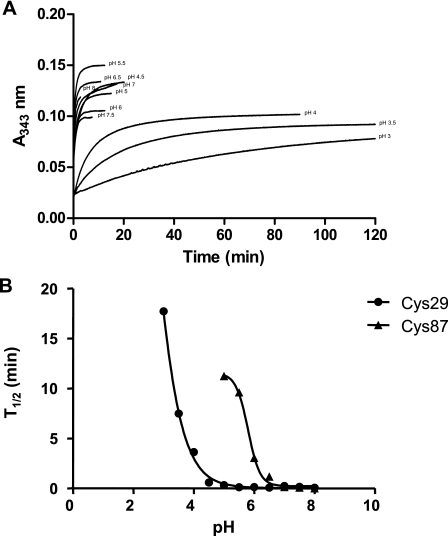
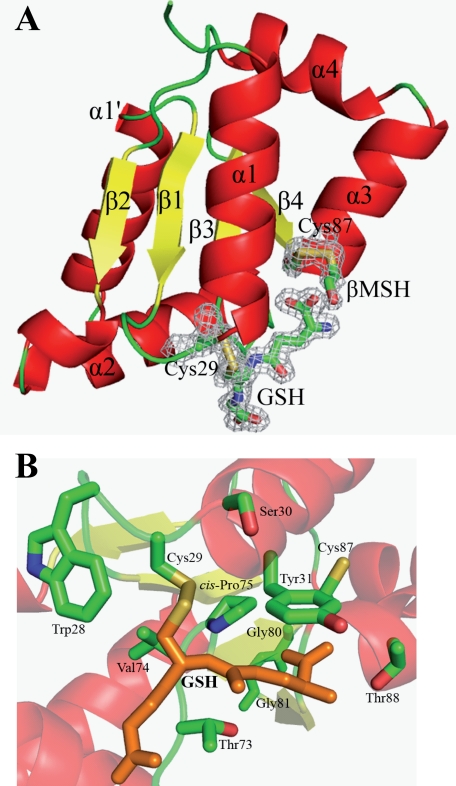
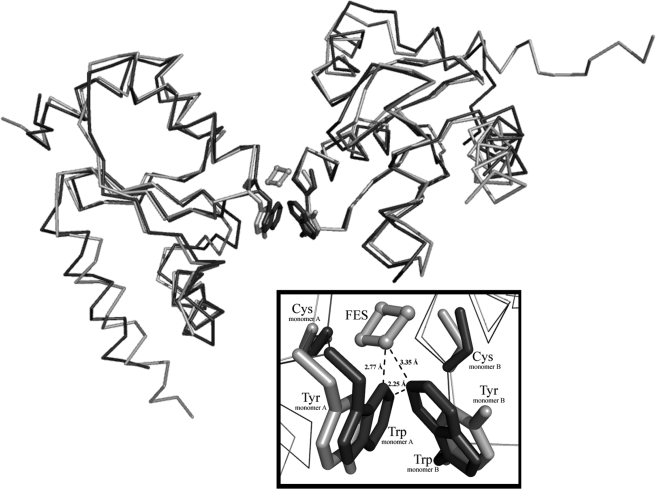
Glutaredoxins (Grxs) are efficient catalysts for the reduction of mixed disulfides in glutathionylated proteins, using glutathione or thioredoxin reductases for their regeneration. Using GFP fusion, we have shown that poplar GrxS12, which possesses a monothiol (28)WCSYS(32) active site, is localized in chloroplasts. In the presence of reduced glutathione, the recombinant protein is able to reduce in vitro substrates, such as hydroxyethyldisulfide and dehydroascorbate, and to regenerate the glutathionylated glyceraldehyde-3-phosphate dehydrogenase. Although the protein possesses two conserved cysteines, it is functioning through a monothiol mechanism, the conserved C terminus cysteine (Cys(87)) being dispensable, since the C87S variant is fully active in all activity assays. Biochemical and crystallographic studies revealed that Cys(87) exhibits a certain reactivity, since its pK(a) is around 5.6. Coupled with thiol titration, fluorescence, and mass spectrometry analyses, the resolution of poplar GrxS12 x-ray crystal structure shows that the only oxidation state is a glutathionylated derivative of the active site cysteine (Cys(29)) and that the enzyme does not form inter- or intramolecular disulfides. Contrary to some plant Grxs, GrxS12 does not incorporate an iron-sulfur cluster in its wild-type form, but when the active site is mutated into YCSYS, it binds a [2Fe-2S] cluster, indicating that the single Trp residue prevents this incorporation.
Figures
References
-
- Bushweller, J. H., Aslund, F., Wuthrich, K., and Holmgren, A. (1992) Biochemistry 31 9288-9293 - PubMed
-
- Rouhier, N., Gelhaye, E., and Jacquot, J. P. (2002) FEBS Lett. 511 145-149 - PubMed
-
- Lillig, C. H., Prior, A., Schwenn, J. D., Aslund, F., Ritz, D., Vlamis-Gardikas, A., and Holmgren, A. (1999) J. Biol. Chem. 74 7695-7698 - PubMed
-
- Johansson, C., Lillig, C. H., and Holmgren, A. (2004) J. Biol. Chem. 279 7537-7543 - PubMed
-
- Tamarit, J., Bellí, G., Cabiscol, E., Herrero, E., and Ros, J. (2003) J. Biol. Chem. 278 25745-25751 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
