

Dissociation of the subunits of the calcium-independent receptor of alpha-latrotoxin as a result of two-step proteolysis
- PMID: 19161337
- PMCID: PMC2682192
- DOI: 10.1021/bi802163p
Dissociation of the subunits of the calcium-independent receptor of alpha-latrotoxin as a result of two-step proteolysis
Abstract
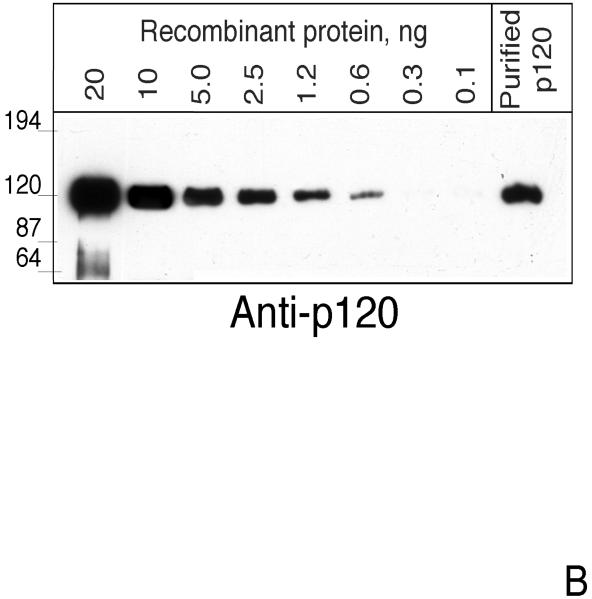
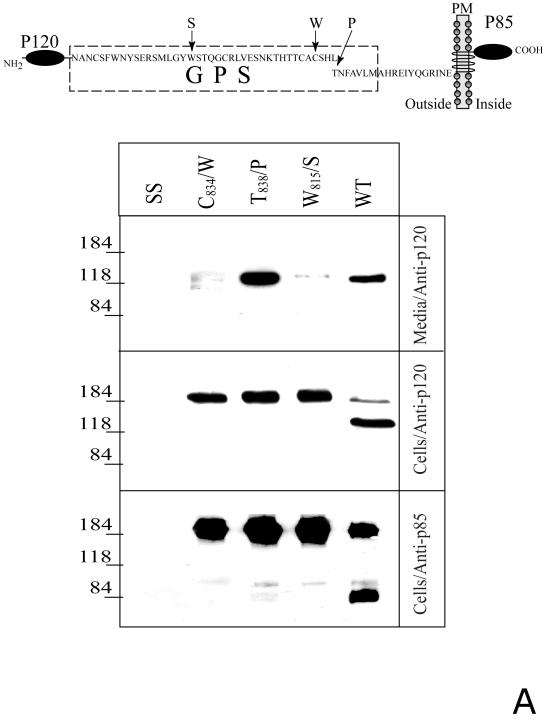
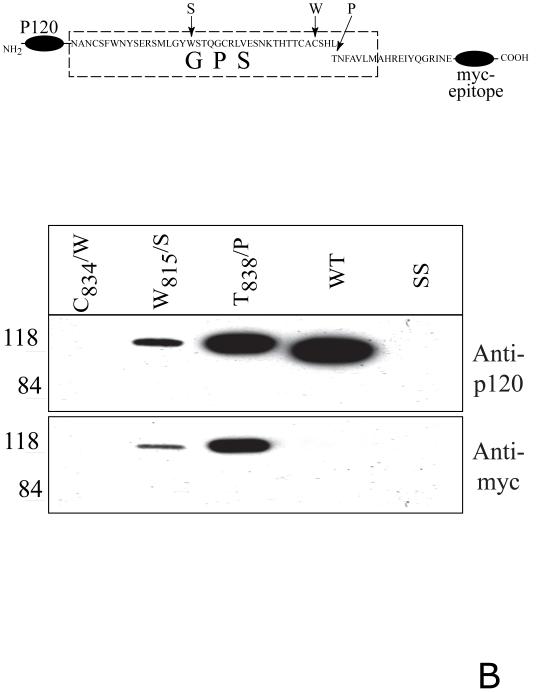
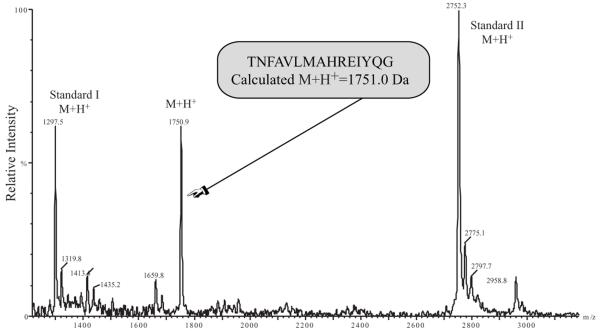
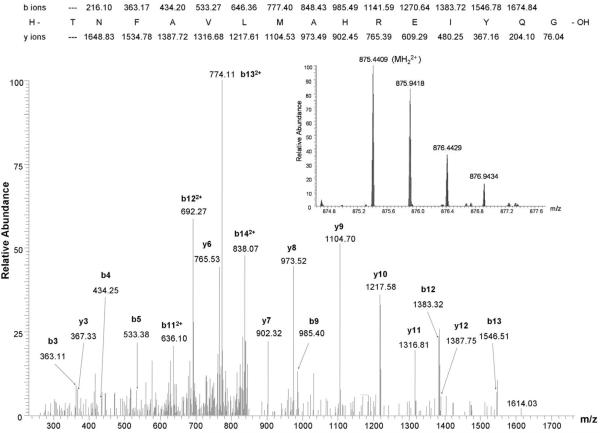
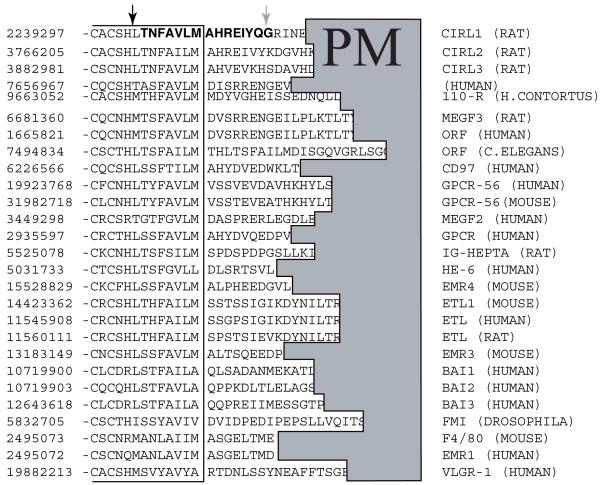
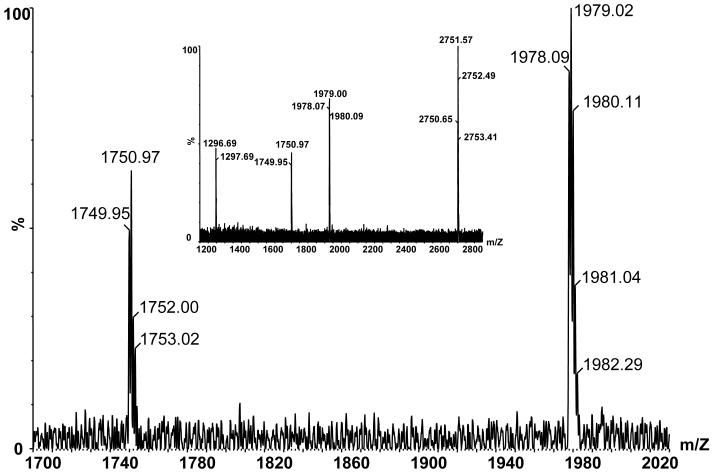
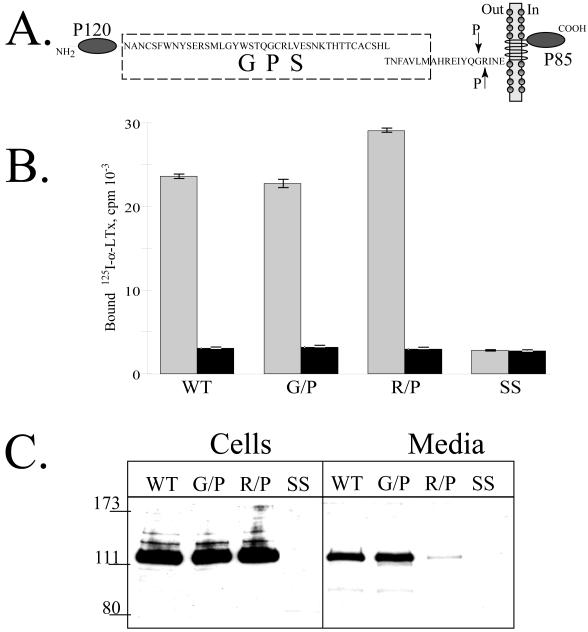
CIRL (the calcium-independent receptor of alpha-latrotoxin), a neuronal cell surface receptor implicated in the regulation of exocytosis, is a member of the GPS family of chimeric cell adhesion/G protein-coupled receptors. The predominant form of CIRL is a membrane-bound complex of two subunits, p120 and p85. Extracellularly oriented p120 contains hydrophilic cell adhesion domains, whereas p85 is a heptahelical membrane protein. Both subunits are encoded by the same gene and represent products of intracellular proteolytic processing of the CIRL precursor. In this study, we demonstrate that a soluble form of CIRL also exists in vitro and in vivo. It results from the further cleavage of CIRL by a second protease. The site of the second cleavage is located in the short N-terminal extracellular tail of p85, between the GPS domain and the first transmembrane segment of CIRL. Thus, the soluble form of CIRL represents a complex of p120 noncovalently bound to a 15 amino acid residue N-terminal peptide fragment of p85. We have previously shown that mutations of CIRL in the GPS domain inhibit intracellular proteolytic processing and also result in the absence of the receptors from the cell surface. Our current data suggest that although CIRL trafficking to the cell membrane is impaired by mutations in the GPS region, it is not blocked completely. However, at the cell surface, the noncleaved mutants are preferentially targeted by the second protease that sheds the extracellular subunit. Therefore, the two-step proteolytic processing may represent a regulatory mechanism that controls cell surface expression of membrane-bound and soluble forms of CIRL.
Figures


References
-
- Hayflick JS. A family of heptahelical receptors with adhesion-like domains: a marriage between two super families. J Recept Signal Transduct Res. 2000;20:119–131. - PubMed
-
- Stacey M, Lin HH, Gordon S, McKnight AJ. LNB-TM7, a group of seven-transmembrane proteins related to family-B G-protein-coupled receptors. Trends in biochemical sciences. 2000;25:284–289. - PubMed
-
- Yona S, Lin HH, Siu WO, Gordon S, Stacey M. Adhesion-GPCRs: emerging roles for novel receptors. Trends in biochemical sciences. 2008;33:491–500. - PubMed
-
- Kwakkenbos MJ, Kop EN, Stacey M, Matmati M, Gordon S, Lin HH, Hamann J. The EGF-TM7 family: a postgenomic view. Immunogenetics. 2004;55:655–666. - PubMed
-
- Usui T, Shima Y, Shimada Y, Hirano S, Burgess RW, Schwarz TL, Takeichi M, Uemura T. Flamingo, a seven-pass transmembrane cadherin, regulates planar cell polarity under the control of Frizzled. Cell. 1999;98:585–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
