

The cerebellum and eye-blink conditioning: learning versus network performance hypotheses
- PMID: 19162131
- PMCID: PMC2822538
- DOI: 10.1016/j.neuroscience.2008.12.042
The cerebellum and eye-blink conditioning: learning versus network performance hypotheses
Abstract
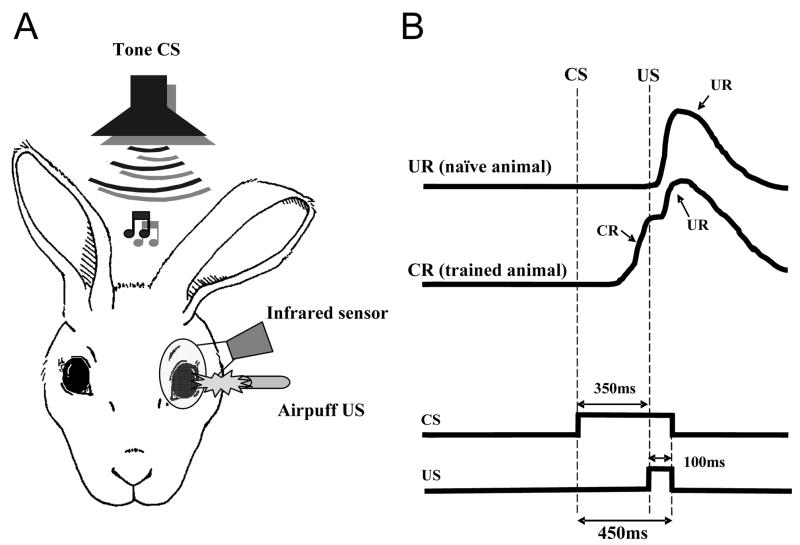
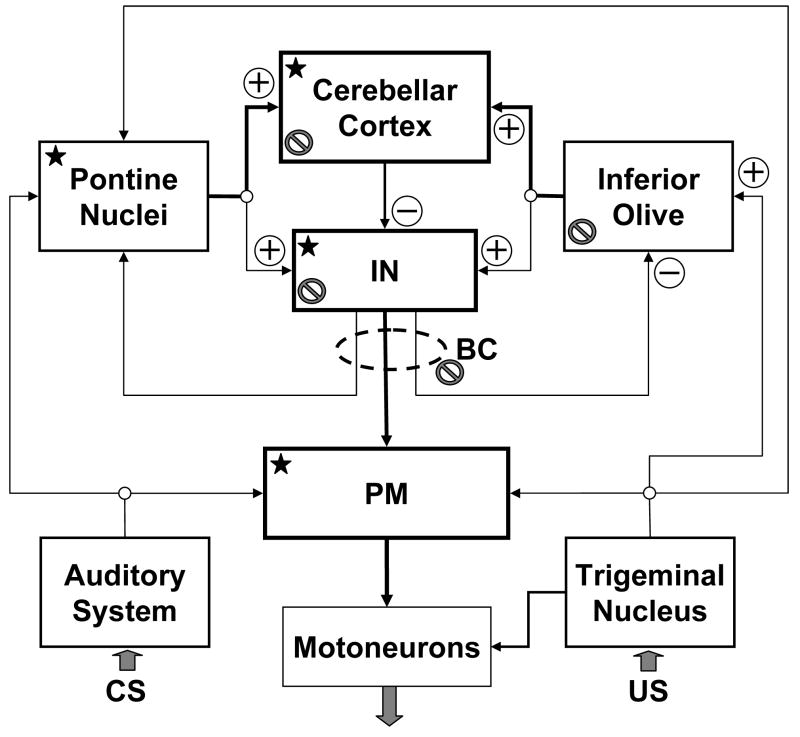
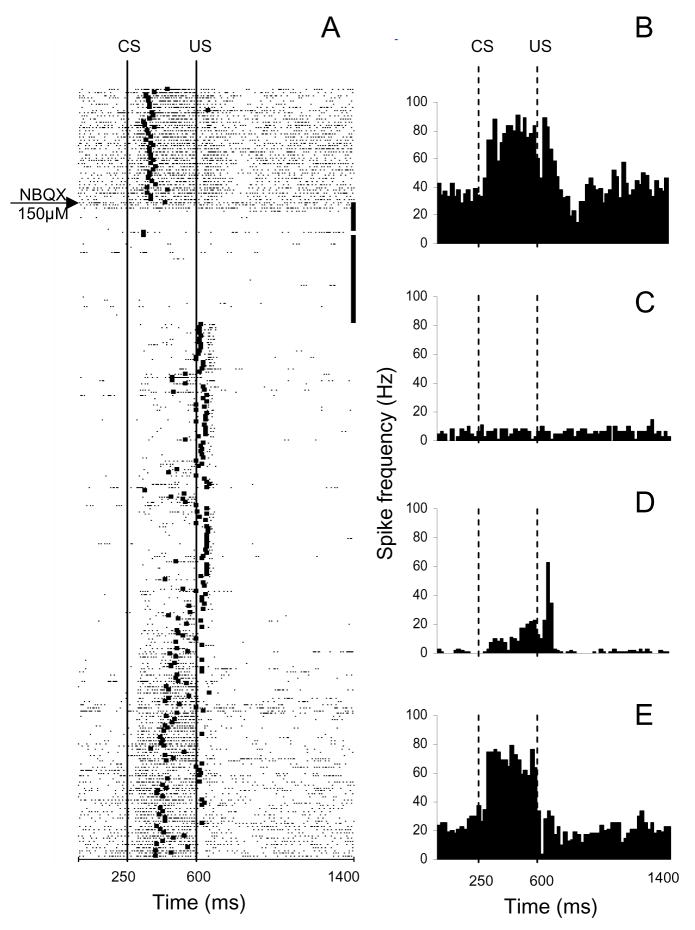
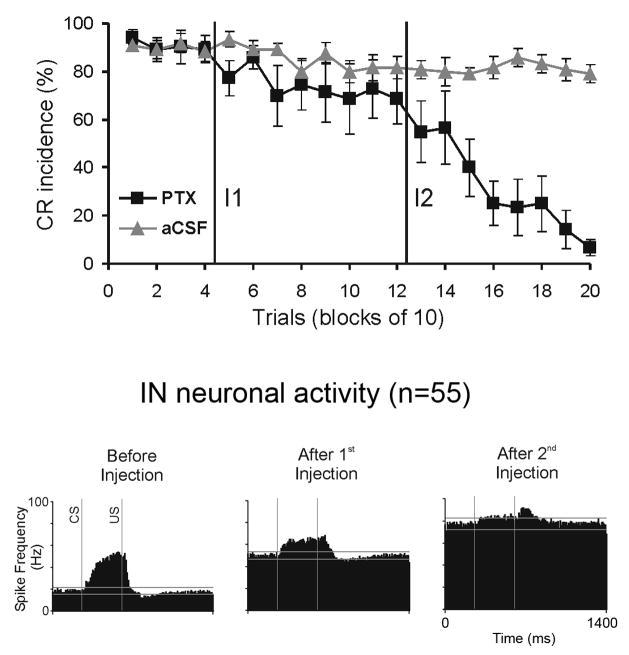
Classical conditioning of the eye-blink reflex in the rabbit is a form of motor learning that is uniquely dependent on the cerebellum. The cerebellar learning hypothesis proposes that plasticity subserving eye-blink conditioning occurs in the cerebellum. The major evidence for this hypothesis originated from studies based on a telecommunications network metaphor of eye-blink circuits. These experiments inactivated parts of cerebellum-related networks during the acquisition and expression of classically conditioned eye blinks in order to determine sites at which the plasticity occurred. However, recent evidence revealed that these manipulations could be explained by a network performance hypothesis which attributes learning deficits to a non-specific tonic dysfunction of eye-blink networks. Since eye-blink conditioning is mediated by a spontaneously active, recurrent neuronal network with strong tonic interactions, differentiating between the cerebellar learning hypothesis and the network performance hypothesis represents a major experimental challenge. A possible solution to this problem is offered by several promising new approaches that minimize the effects of experimental interventions on spontaneous neuronal activity. Results from these studies indicate that plastic changes underlying eye-blink conditioning are distributed across several cerebellar and extra-cerebellar regions. Specific input interactions that induce these plastic changes as well as their cellular mechanisms remain unresolved.
Figures
References
-
- Aksenov D, Serdyukova N, Irwin K, Bracha V. GABA neurotransmission in the cerebellar interposed nuclei: involvement in classically conditioned eyeblinks and neuronal activity. J Neurophysiol. 2004;91:719–727. - PubMed
-
- Aksenov D, Serdyukova N, Bloedel JR, Bracha V. 2004 Abstract Viewer/Itinerary Planner. Washington, DC: Society for Neuroscience; 2004. The role of tonic activity and phasic modulation of interposed cerebellar nuclear cells in the expression of conditioned eyeblinks. 136.6.
-
- Albus JS. A theory of cerebellar function. Math Biosci. 1971;10:25–61.
-
- Attwell PJ, Cooke SF, Yeo CH. Cerebellar function in consolidation of a motor memory. Neuron. 2002a;34:1011–1020. - PubMed
-
- Attwell PJ, Ivarsson M, Millar L, Yeo CH. Cerebellar mechanisms in eyeblink conditioning. Ann N Y Acad Sci. 2002b;978:79–92. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
