

Seminal fluid drives expansion of the CD4+CD25+ T regulatory cell pool and induces tolerance to paternal alloantigens in mice
- PMID: 19164169
- PMCID: PMC2849830
- DOI: 10.1095/biolreprod.108.074658
Seminal fluid drives expansion of the CD4+CD25+ T regulatory cell pool and induces tolerance to paternal alloantigens in mice
Abstract
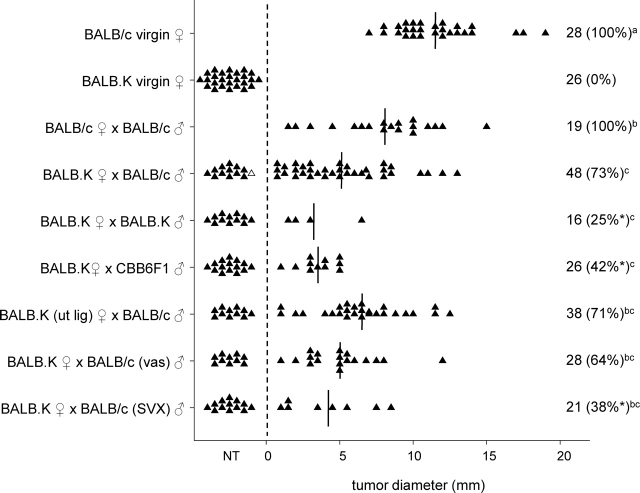
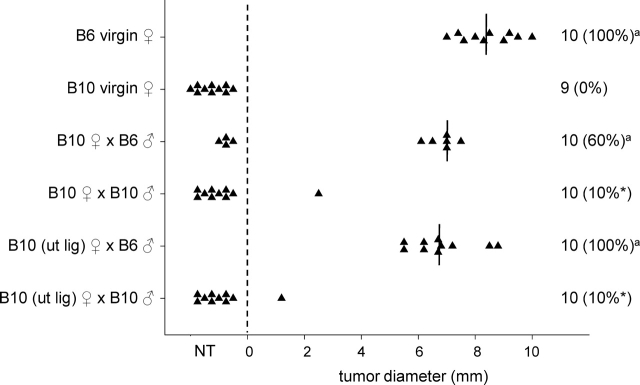
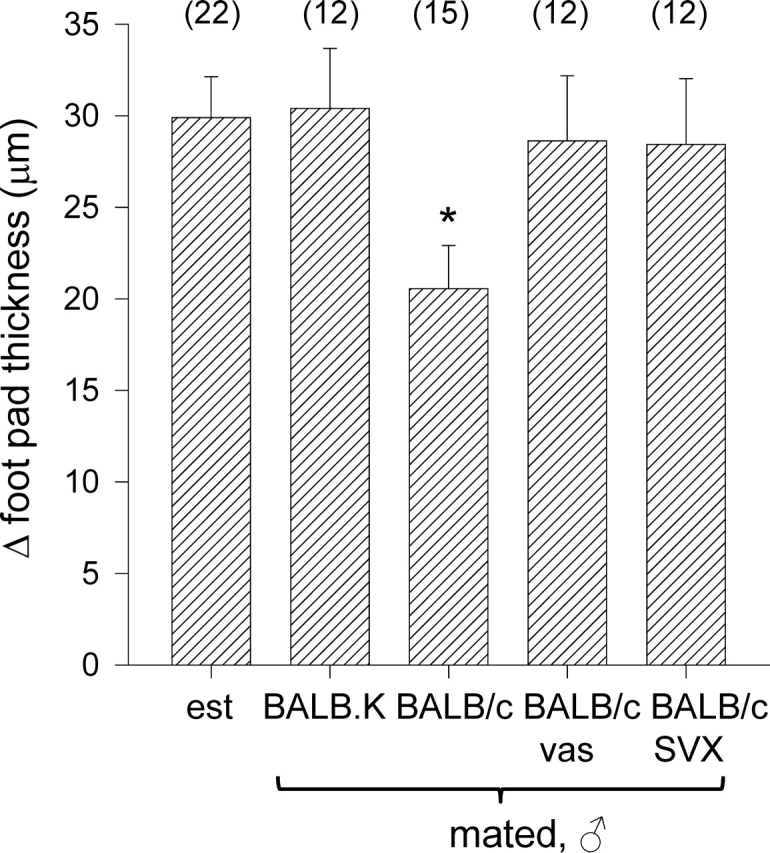
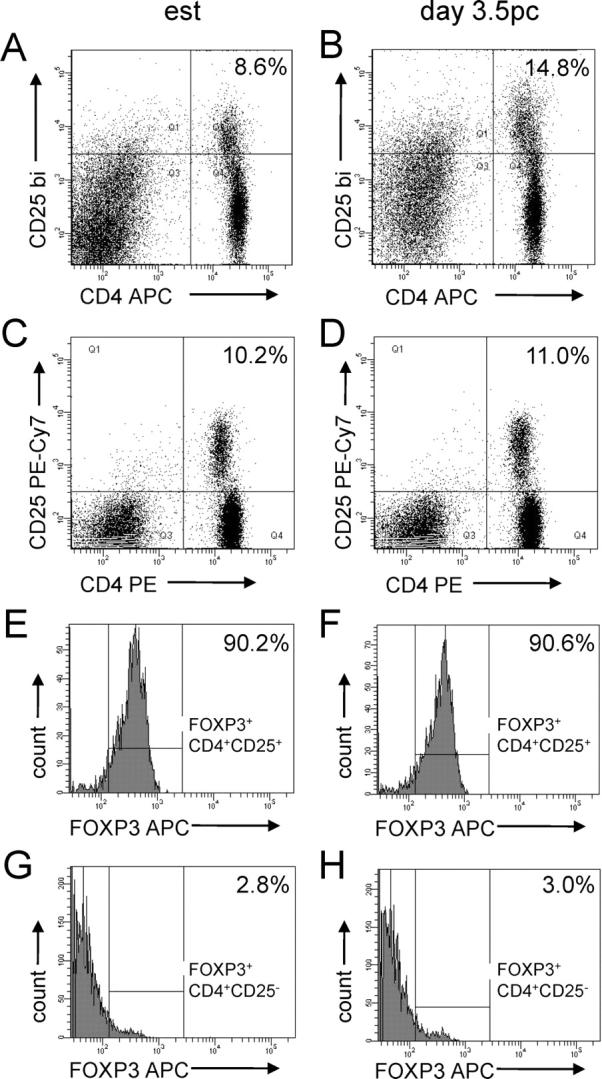
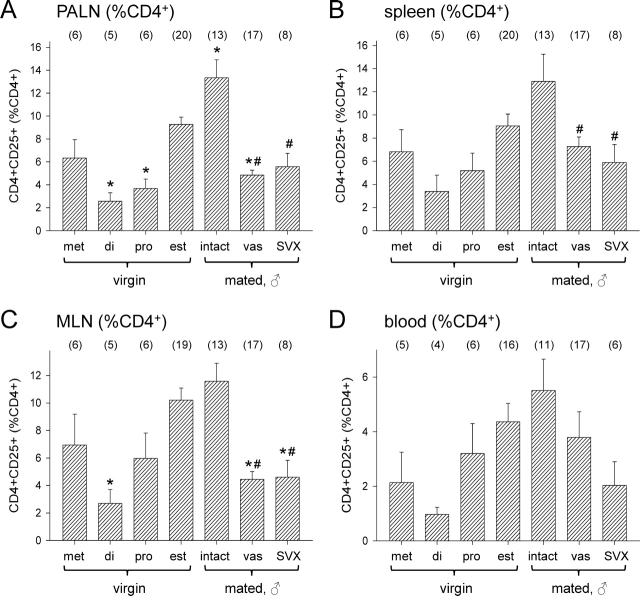
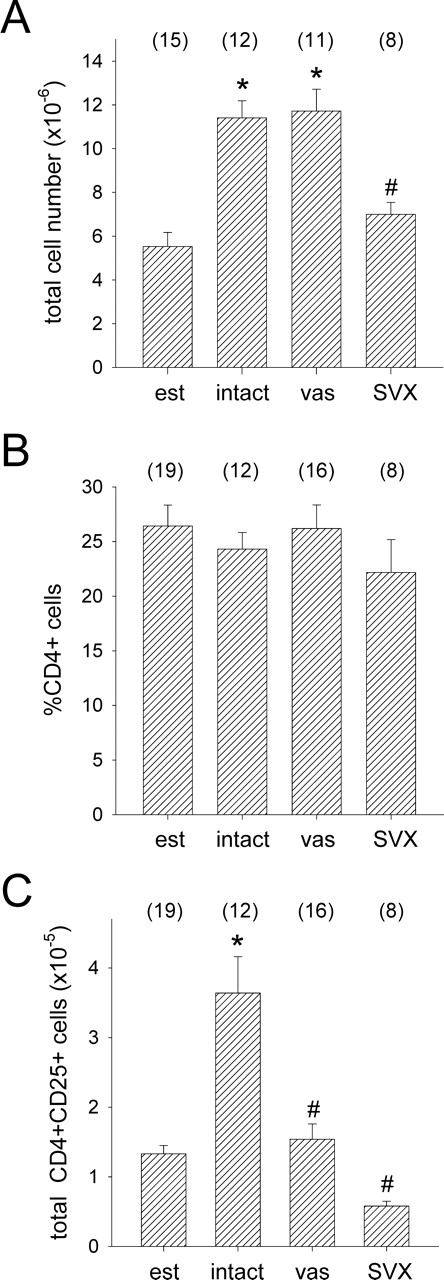
T regulatory (Treg) cells are implicated in maternal immune tolerance of the conceptus at implantation; however, the antigenic and regulatory signals controlling Treg cells in early pregnancy are undefined. To examine the role of male seminal fluid in tolerance induction, the effect of exposure to seminal fluid at mating on responsiveness to paternal alloantigens was examined using paternal tumor cell grafts and by delayed-type hypersensitivity (DTH) challenge on Day 3.5 postcoitum. Exposure to seminal fluid inhibited rejection of paternal tumor cells, independently of fertilization and embryo development, while seminal fluid from major histocompatability complex (MHC)-dissimilar males was less effective. Similarly, mating with intact males suppressed the DTH response to paternal alloantigens in an MHC-specific fashion. Excision of the seminal vesicle glands diminished the tolerance-inducing activity of seminal fluid. Mating with intact males caused an increase in CD4(+)CD25(+) cells expressing FOXP3 in the para-aortic lymph nodes draining the uterus, beyond the estrus-associated peak in cycling mice. The increase in CD4(+)CD25(+) cells was abrogated when males were vasectomized or seminal vesicles were excised. Collectively, these data provide evidence that exposure to seminal fluid at mating promotes a state of functional tolerance to paternal alloantigens that may facilitate maternal acceptance of the conceptus at implantation, and the effects of seminal fluid are likely to be mediated by expansion of the Treg cell pool. Both seminal plasma and sperm components of the seminal fluid are necessary to confer full tolerance and elicit the Treg cell response, potentially through provision of immune-deviating cytokines and antigens, respectively.
Figures
References
-
- Trowsdale J, Betz AG.Mother's little helpers: mechanisms of maternal-fetal tolerance. Nat Immunol 2006; 7: 241–246. - PubMed
-
- Guleria I, Sayegh MH.Maternal acceptance of the fetus: true human tolerance. J Immunol 2007; 178: 3345–3351. - PubMed
-
- Tafuri A, Alferink J, Moller P, Hammerling GJ, Arnold B.T cell awareness of paternal alloantigens during pregnancy. Science 1995; 270: 630–633. - PubMed
-
- Jiang SP, Vacchio MS.Multiple mechanisms of peripheral T cell tolerance to the fetal “allograft.” J Immunol 1998; 160: 3086–3090. - PubMed
-
- Zhou M, Mellor AL.Expanded cohorts of maternal CD8+ T-cells specific for paternal MHC class I accumulate during pregnancy. J Reprod Immunol 1998; 40: 47–62. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
