

Susceptibility to autoimmunity and B cell resistance to apoptosis in mice lacking androgen receptor in B cells
- PMID: 19164450
- PMCID: PMC2667704
- DOI: 10.1210/me.2008-0106
Susceptibility to autoimmunity and B cell resistance to apoptosis in mice lacking androgen receptor in B cells
Erratum in
-
Corrigendum to: "Susceptibility to Autoimmunity and B Cell Resistance to Apoptosis in Mice Lacking Androgen Receptor in B Cells".Endocrinology. 2020 Sep 1;161(9):bqaa118. doi: 10.1210/endocr/bqaa118. Endocrinology. 2020. PMID: 32822479 No abstract available.
Abstract
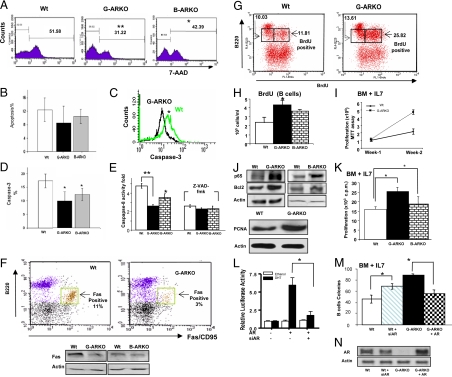
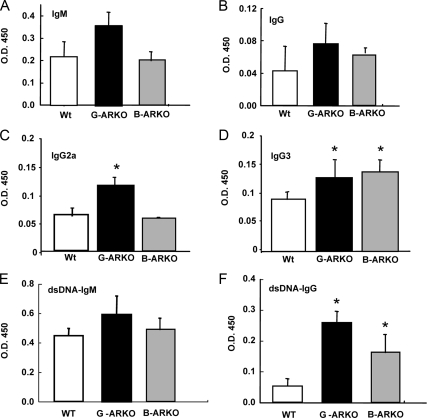
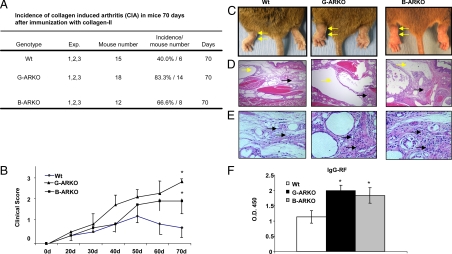
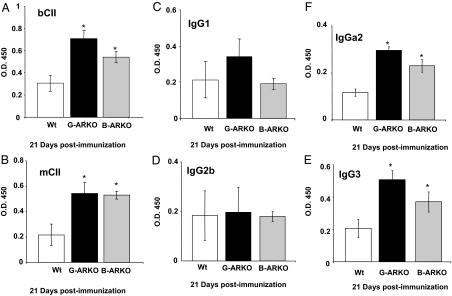
Estrogens have been linked to a higher female incidence of autoimmune diseases. The role of androgen and the androgen receptor (AR) in autoimmune diseases, however, remains unclear. Here we report that the lack of AR in B cells in different strains of mice, namely general AR knockout, B cell-specific AR knockout, and naturally occurring testicular feminization mutation AR-mutant mice, as well as castrated wild-type mice, results in increased B cells in blood and bone marrow. Analysis of the targeted mice, together with bone marrow transplantation using Rag1(-/-) recipients, overexpression of retrovirally encoded AR-cDNA, and small interfering RNA-mediated AR mRNA knockdown approaches also show that the B cell expansion results from resistance to apoptosis and increased proliferation of bone marrow precursor B cells, accompanied by changes in several key modulators related to apoptosis, such as Fas/FasL signals, caspases-3/-8, nuclear factor-kappaB, and Bcl-2. We also show that the effects of AR loss are, in part, B cell intrinsic. Mice bearing AR-deficient B cells show increased levels of serum IgG2a and IgG3 as well as basal double-stranded DNA-IgG antibodies and are more vulnerable to development of collagen-induced arthritis. Together, these data indicate that androgen/AR play a crucial role in B cell homeostasis and tolerance. Therapies targeting AR might provide an alternative strategy with which to battle autoimmune diseases.
Figures
References
-
- Chang CS, Kokontis J, Liao ST 1988 Molecular cloning of human and rat complementary DNA encoding androgen receptors. Science 240:324–326 - PubMed
-
- Heinlein CA, Chang C 2004 Androgen receptor in prostate cancer. Endocr Rev 25:276–308 - PubMed
-
- Tsai MJ, O'Malley BW 1994 Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu Rev Biochem 63:451–486 - PubMed
-
- Rochette-Egly C 2005 Dynamic combinatorial networks in nuclear receptor-mediated transcription. J Biol Chem 280:32565–32568 - PubMed
-
- Abu EO, Horner A, Kusec V, Triffitt JT, Compston JE 1997 The localization of androgen receptors in human bone. J Clin Endocrinol Metab 82:3493–3497 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
