

Substrate selectivity of 5-hydroxyeicosanoid dehydrogenase and its inhibition by 5-hydroxy-Delta6-long-chain fatty acids
- PMID: 19164464
- PMCID: PMC2670600
- DOI: 10.1124/jpet.108.143453
Substrate selectivity of 5-hydroxyeicosanoid dehydrogenase and its inhibition by 5-hydroxy-Delta6-long-chain fatty acids
Abstract
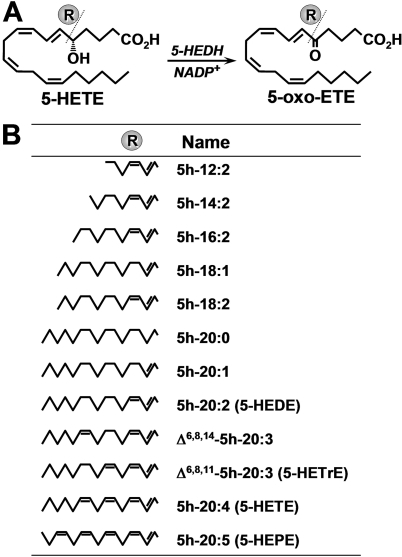
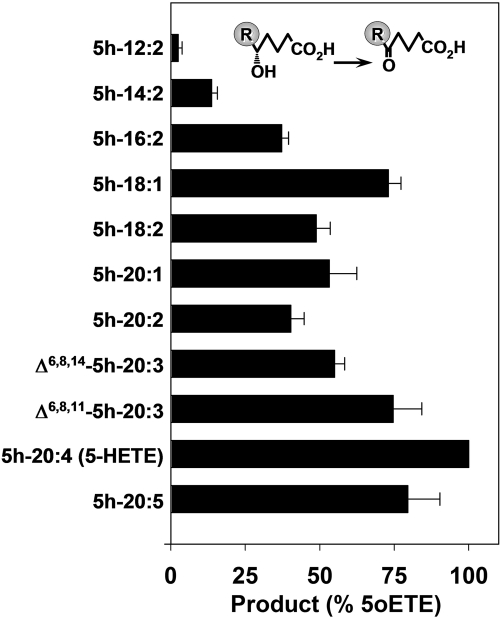
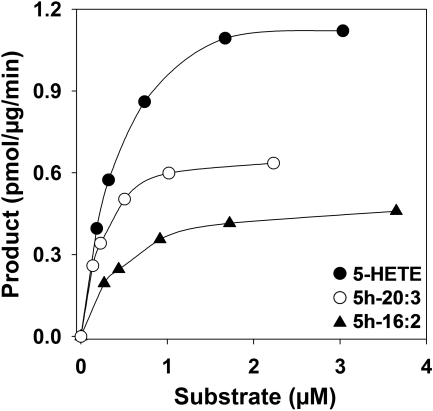
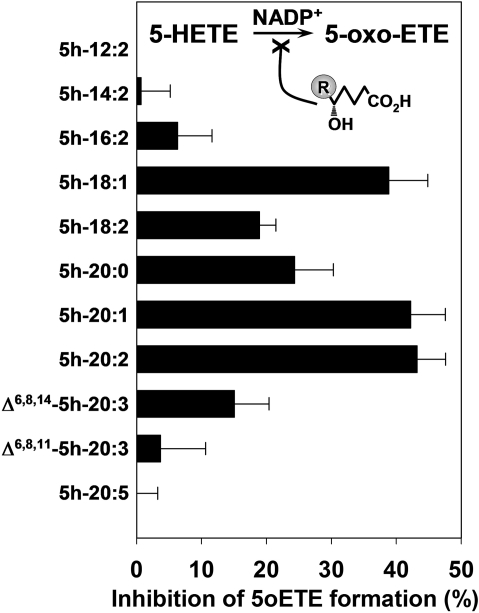
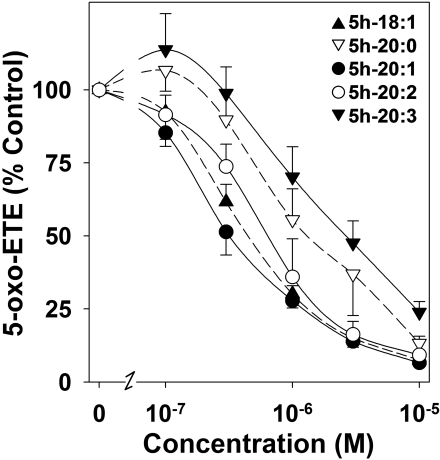
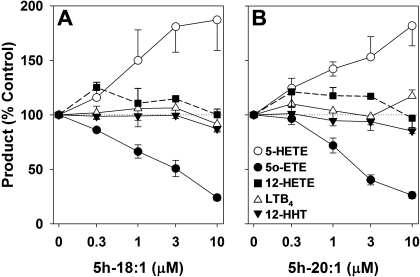
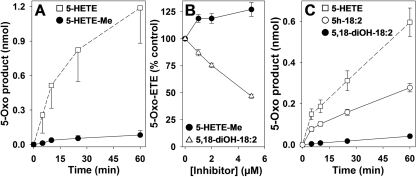
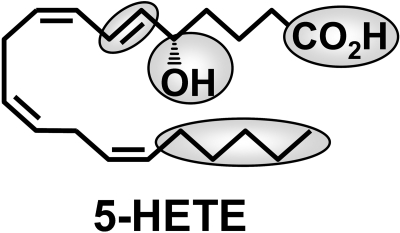
5-Oxo-6E,8Z,11Z,14Z-eicosatetraenoic acid (5-oxo-ETE) is a metabolite of the 5-lipoxygenase (5-LO) product 5S-hydroxy-6E,8Z,11Z,14Z-eicosatetraenoic acid (5-HETE), formed by the microsomal enzyme 5-hydroxyeicosanoid dehydrogenase (5-HEDH). 5-oxo-ETE is a chemoattractant for neutrophils and eosinophils, both in vitro and in vivo. To examine the substrate selectivity of 5-HEDH and to search for potential inhibitors, we prepared a series of 5S-hydroxy fatty acids (C(12) to C(20) containing zero to four double bonds) by total chemical synthesis and examined their metabolism by microsomes from monocytic U937 cells. Although most of these fatty acids were oxidized to their 5-oxo metabolites by 5-HEDH, 5-HETE seemed to be the best substrate. However, substrates containing less than 16 carbons, a methylated alpha-carboxyl group, or a hydroxyl group at the omega-end of the molecule were not substantially metabolized. Some of the fatty acids tested were fairly potent inhibitors of the formation of 5-oxo-ETE by 5-HEDH, in particular 5-hydroxy-6-octadecenoic acid and 5-hydroxy-6-eicosenoic acid. Both substances selectively inhibited 5-oxo-ETE formation by human peripheral blood mononuclear cells incubated with arachidonic acid and calcium ionophore without affecting the formation of leukotriene B(4), 12-HETE, or 12-hydroxy-5,8,10-heptadecatrienoic acid. We conclude that the requirements for appreciable metabolism by 5-HEDH include a chain length of at least 16 carbons, a free alpha-carboxyl group, and a hydrophobic group at the omega-end of the molecule. 5-Hydroxy-Delta(6) C(18) and C(20) fatty acids selectively inhibit 5-HEDH without inhibiting 5-LO, leukotriene A(4) hydrolase, 12-lipoxygenase, or cyclooxygenase. Such compounds may be useful in defining the role of 5-oxo-ETE and its mechanism of synthesis.
Figures
References
-
- Dallaire MJ, Ferland C, Pagé N, Lavigne S, Davoine F, and Laviolette M (2003) Endothelial cells modulate eosinophil surface markers and mediator release. Eur Respir J 21 918-924. - PubMed
-
- Erlemann KR, Rokach J, and Powell WS (2004) Oxidative stress stimulates the synthesis of the eosinophil chemoattractant 5-oxo-6,8,11,14-eicosatetraenoic acid by inflammatory cells. J Biol Chem 279 40376-40384. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
