

Characterization of the thymic IL-7 niche in vivo
- PMID: 19164539
- PMCID: PMC2635771
- DOI: 10.1073/pnas.0809559106
Characterization of the thymic IL-7 niche in vivo
Abstract
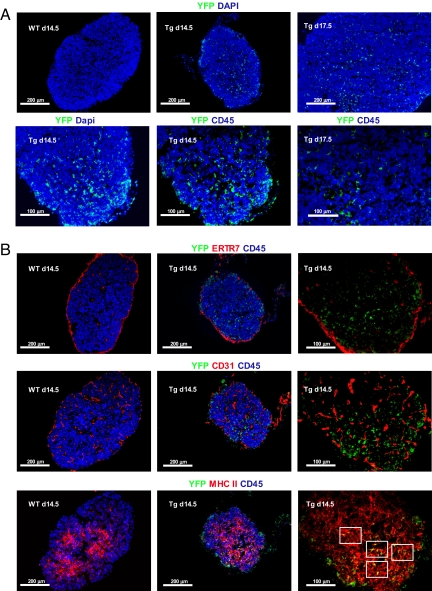
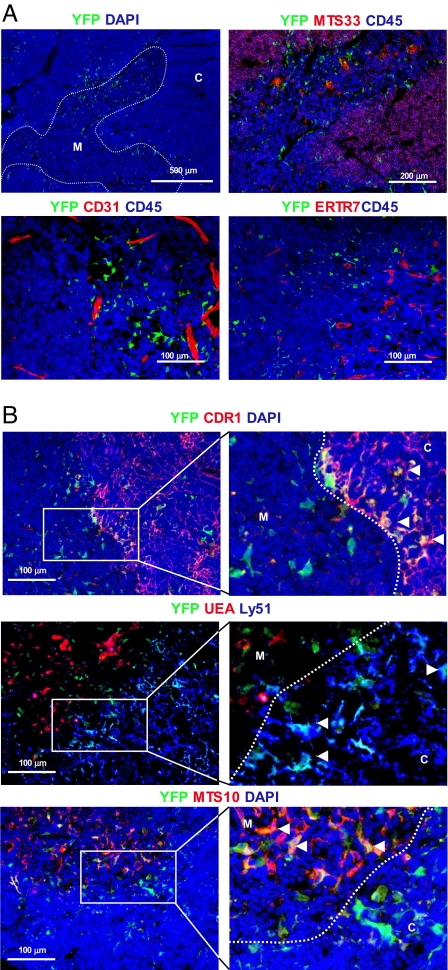
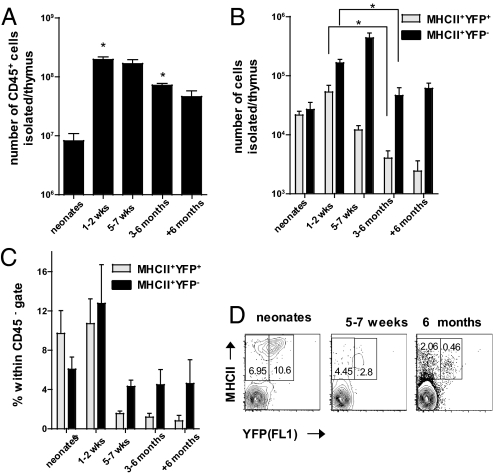
The thymus represents the "cradle" for T cell development, with thymic stroma providing multiple soluble and membrane cues to developing thymocytes. Although IL-7 is recognized as an essential factor for thymopoiesis, the "environmental niche" of thymic IL-7 activity remains poorly characterized in vivo. Using bacterial artificial chromosome transgenic mice in which YFP is under control of IL-7 promoter, we identify a subset of thymic epithelial cells (TECs) that co-express YFP and high levels of Il7 transcripts (IL-7(hi) cells). IL-7(hi) TECs arise during early fetal development, persist throughout life, and co-express homeostatic chemokines (Ccl19, Ccl25, Cxcl12) and cytokines (Il15) that are critical for normal thymopoiesis. In the adult thymus, IL-7(hi) cells localize to the cortico-medullary junction and display traits of both cortical and medullary TECs. Interestingly, the frequency of IL-7(hi) cells decreases with age, suggesting a mechanism for the age-related thymic involution that is associated with declining IL-7 levels. Our temporal-spatial analysis of IL-7-producing cells in the thymus in vivo suggests that thymic IL-7 levels are dynamically regulated under distinct physiological conditions. This IL-7 reporter mouse provides a valuable tool to further dissect the mechanisms that govern thymic IL-7 expression in vivo.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Jameson SC, Hogquist KA, Bevan MJ. Positive selection of thymocytes. Annu Rev Immunol. 1995;13:93–126. - PubMed
-
- Kyewski B, Klein L. A central role for central tolerance. Annu Rev Immunol. 2006;24:571–606. - PubMed
-
- Anderson G, Moore NC, Owen JJ, Jenkinson EJ. Cellular interactions in thymocyte development. Annu Rev Immunol. 1996;14:73–99. - PubMed
-
- Derbinski J, Schulte A, Kyewski B, Klein L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat Immunol. 2001;2:1032–1039. - PubMed
-
- Petrie HT, Zuniga-Pflucker JC. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu Rev Immunol. 2007;25:649–679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
