

Induction of TLR4-target genes entails calcium/calmodulin-dependent regulation of chromatin remodeling
- PMID: 19164553
- PMCID: PMC2633529
- DOI: 10.1073/pnas.0811274106
Induction of TLR4-target genes entails calcium/calmodulin-dependent regulation of chromatin remodeling
Abstract
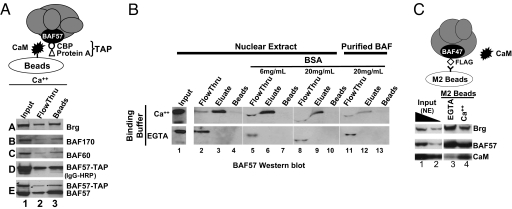
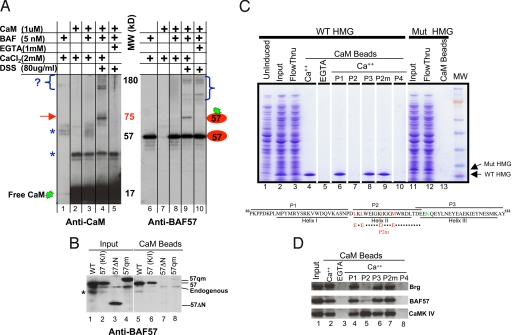
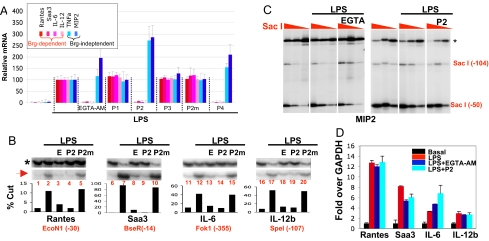
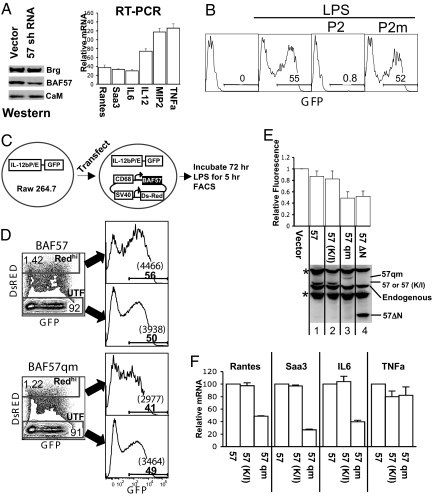
Upon toll-like receptor 4 (TLR4) signaling in macrophages, the mammalian Swi/Snf-like BAF chromatin remodeling complex is recruited to many TLR4 target genes where it remodels their chromatin to promote transcription. Here, we show that, surprisingly, recruitment is not sufficient for chromatin remodeling; a second event, dependent on calcium/calmodulin (CaM), is additionally required. Calcium/CaM directly binds the HMG domain of the BAF57 subunit within the BAF complex. Calcium/CaM antagonists, including a CaM-binding peptide derived from BAF57, abolish BAF-dependent remodeling and gene expression without compromising BAF recruitment. BAF57 RNAi and BAF57 dominant negative mutants defective in CaM binding similarly impair the induction of BAF target genes. Our data implicate calcium/CaM in TLR4 signaling, and reveal a previously undescribed, recruitment-independent mode of regulation of the BAF complex that is probably achieved through a direct CaM-BAF interaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Crystal structure of the HMG domain of human BAF57 and its interaction with four-way junction DNA.Biochem Biophys Res Commun. 2020 Dec 17;533(4):919-924. doi: 10.1016/j.bbrc.2020.09.094. Epub 2020 Sep 30. Biochem Biophys Res Commun. 2020. PMID: 33010889
-
Molecular basis of CD4 repression by the Swi/Snf-like BAF chromatin remodeling complex.Eur J Immunol. 2009 Feb;39(2):580-8. doi: 10.1002/eji.200838909. Eur J Immunol. 2009. PMID: 19180471 Free PMC article.
-
Reciprocal regulation of CD4/CD8 expression by SWI/SNF-like BAF complexes.Nature. 2002 Jul 11;418(6894):195-9. doi: 10.1038/nature00876. Nature. 2002. PMID: 12110891
-
The developmental and pathogenic roles of BAF57, a special subunit of the BAF chromatin-remodeling complex.FEBS Lett. 2016 Jun;590(11):1555-69. doi: 10.1002/1873-3468.12201. Epub 2016 May 20. FEBS Lett. 2016. PMID: 27149204 Review.
-
BAFfling pathologies: Alterations of BAF complexes in cancer.Cancer Lett. 2018 Apr 10;419:266-279. doi: 10.1016/j.canlet.2018.01.046. Epub 2018 Jan 31. Cancer Lett. 2018. PMID: 29374542 Review.
Cited by
-
Critical role for calcium mobilization in activation of the NLRP3 inflammasome.Proc Natl Acad Sci U S A. 2012 Jul 10;109(28):11282-7. doi: 10.1073/pnas.1117765109. Epub 2012 Jun 25. Proc Natl Acad Sci U S A. 2012. PMID: 22733741 Free PMC article.
-
MT1-MMP regulates the PI3Kδ·Mi-2/NuRD-dependent control of macrophage immune function.Genes Dev. 2012 Feb 15;26(4):395-413. doi: 10.1101/gad.178749.111. Genes Dev. 2012. PMID: 22345520 Free PMC article.
-
Dynamic BRG1 recruitment during T helper differentiation and activation reveals distal regulatory elements.Mol Cell Biol. 2011 Apr;31(7):1512-27. doi: 10.1128/MCB.00920-10. Epub 2011 Jan 24. Mol Cell Biol. 2011. PMID: 21262765 Free PMC article.
-
Oocyte activation is a cytoplasm-confined event so far: what about the nucleus?Reproduction. 2024 Feb 2;167(3):e230360. doi: 10.1530/REP-23-0360. Print 2024 Mar 1. Reproduction. 2024. PMID: 38112585 Free PMC article.
-
The immunomodulatory role of matrix metalloproteinases in colitis-associated cancer.Front Immunol. 2023 Jan 19;13:1093990. doi: 10.3389/fimmu.2022.1093990. eCollection 2022. Front Immunol. 2023. PMID: 36776395 Free PMC article. Review.
References
-
- Chi T. A BAF-centred view of the immune system. Nat Rev Immunol. 2004;4:965–977. - PubMed
-
- Agalioti T, et al. Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell. 2000;103:667–678. - PubMed
-
- Phelan ML, Sif S, Narlikar GJ, Kingston RE. Reconstitution of a core chromatin remodeling complex from SWI/SNF subunits. Mol Cell. 1999;3:247–253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
