

doi: 10.1073/pnas.0809192106.
Epub 2009 Jan 21.
Intersubunit allosteric communication mediated by a conserved loop in the MCM helicase
Affiliations
- PMID: 19164574
- PMCID: PMC2633543
- DOI: 10.1073/pnas.0809192106
Item in Clipboard
Intersubunit allosteric communication mediated by a conserved loop in the MCM helicase
Proc Natl Acad Sci U S A.
.
Abstract
The minichromosome maintenance (MCM) helicase is the presumptive replicative helicase in archaea and eukaryotes. The archaeal homomultimeric MCM has a two-tier structure. One tier contains the AAA+ motor domains of the proteins, and these are the minimal functional helicase domains. The second tier is formed by the N-terminal domains. These domains are not essential for MCM helicase activity but act to enhance the processivity of the helicase. We reveal that a conserved loop facilitates communication between processivity and motor tiers. Interestingly, this allostery seems to be mediated by interactions between, rather than within, individual protomers in the MCM ring.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

A conserved loop in the N-terminal tier of MCM is required for intersubunit communication. (A) Mutant doping assay using the C-half of both wild-type MCM (residues 267–612) and Walker A lysine–alanine mutant (K346A in the full-length protein) in varying proportions in helicase assays. The red line indicates the nonlinear behavior of the full-length proteins (ref. 8). (B) Structure of the N-terminal 265 residues of SsoMCM [Protein Data Bank (PDB) ID code 2vl6]. The figure was by using PyMOL (http:://pymol.sourceforge.net ). The ACL is highlighted in red, and residues E202, E203, and Q208 are shown in brown stick form. The N-terminal DNA-binding β-hairpin is shown in dark blue. A zinc ion is shown as a brown sphere. (C) Sequence line-up of the ACL of seven representative archaeal MCM sequences (Afu, Archaeoglogus fulgidus; Csy, Cenarchaeum symbiosum; Hby, Hyperthermus butylicus; Mth, M. thermautotrophicus; Neq, Nanoarchaeum equitans, Sso, S. solfataricus; Tpe, Thermofilum pendens) together with human MCM2–9. The ACL is indicated in red, and residues corresponding to SsoMCM E202, E203, and Q208 are indicated by brown asterisks. (D) DNA binding to a flayed duplex substrate for wild-type MCM (WT-MCM), ΔACL, and ACL-PM (E202A, E203A, and Q207A) are shown (analogous results for ssDNA and dsDNA are shown in Fig S2 ). Error bars in this and all other panels are ±SD. (E) DNA helicase activity of WT, ΔACL, and ACL-PM MCM. (F) ATPase rates of the WT, ΔACL, and ACL-PM MCM. Values are picomoles of ATP hydrolyzed per second per nanomole of MCM (as monomer). (G) Mutant doping with ΔACL or Walker A mutant (K346A) into wild-type MCM. The helicase activities of the protein mixtures were measured and plotted relative to the activity of wild-type protein. Simulations (red and cyan lines) were performed as described in ref. .

Functional consequences of ACL deletion. (A) Pulldown assays of wild-type (WT) or ΔACL versions of the N-terminal domains of MCM (residues 1–266) on a matrix with immobilized C-half of MCM. After pulldown and washing, proteins were eluted by boiling in SDS/PAGE loading buffer. Proteins were detected after SDS/PAGE by staining with Coomassie brilliant blue. The positions of the various species are indicated. Analogous assays performed in the presence of ADP, ATP, AMP-PNP, and ADP-AlFx are shown in Fig. S5 . (B) Helicase assays mediated by 0.5 μM C-half of MCM supplemented with increasing amounts of either wild-type or ΔACL N-half. (C) Helicase assays with full-length proteins with either wild-type, deleted (ΔNβHP) or point-mutated (KR246AA) N-terminal β-hairpin proteins with additional deletion of the ACL (ΔACL) as indicated. (D) Comparison of the relative processivity of wild-type or ΔNβHPΔACL proteins.

Nucleotide-modifiable communication between the C-terminal pre-Sensor 1 β-hairpin and the ACL. (A) (Left) Superposition of the MthMCM EM structure (4) with the known structure of MthMCM N-terminal domains (in cyan; the ACL is highlighted in red). The C-terminal domains are modeled based on the structure of a bacterial ATPase (PDB ID code 2R44) that is predicted to have secondary structure architecture similar to MCM (conserved domain database expected value = 1×e−4). (Right) Enlargement of the selected area. The C-terminal β-hairpin is highlighted in yellow. (B) DEER time trace after background correction shown in black for nucleotide-free MCM and in red MCM in the presence of 10 mM ATP. The best-fit line was found by using a Tikhonov regularization parameter of 1,000 and is shown in red. (C) The distance distributions corresponding to the DEER traces shown in B; black is nucleotide-free MCM, and red is MCM in the presence of 10 mM ATP. (D) Results of cross-linking assays with S206C (Ncys), K430C (Csys) single mutants or S206CK430C (NcysCsys) double mutants. The positions of monomer, dimer, and higher-order species are indicated. Wild-type (cysteine-free protein) is labeled W. M refers to mock-treated (reaction minus BMOE), and X indicates reactions treated with BMOE. (E) Cross-linking assays performed between S206C (Ncys) and K430C (Ccys) in the presence or absence of the indicated nucleotide (at 10 mM).
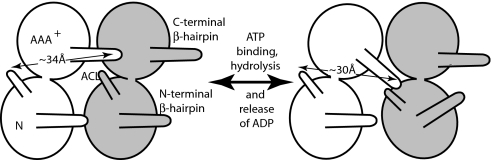
Model for the interplay between C-terminal β-hairpin of one subunit and the ACL of its neighboring subunit in modulation of the position of the neighbor's N-terminal β-hairpin. We propose that, during the cycle of ATP binding and hydrolysis and release of ADP from one subunit, its C-terminal β-hairpin is repositioned, allowing it to contact the ACL of its neighbor and thus reposition the N-terminal β-hairpin of that subunit. (Left) The 34-Å distance indicated in the white subunit would correspond to the ATP-bound version of the protein. (Right) The 30-Å distance between the white subunit ACL and C-terminal β-hairpin and close apposition of the β-hairpin to the ACL of the neighboring subunit would correspond to the nucleotide-free form.
References
-
- Costa A, Onesti S. The MCM complex: (Just) a replicative helicase? Biochem Soc Trans. 2008;36:136–140. - PubMed
-
- Fletcher RJ, et al. The structure and function of MCM from archaeal M. thermoautotrophicum. Nat Struct Biol. 2003;10:160–167. - PubMed
-
- McGeoch AT, Trakselis MA, Laskey RA, Bell SD. Organization of the archaeal MCM complex on DNA and implications for the helicase mechanism. Nat Struct Mol Biol. 2005;12:756–762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
