

Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis
- PMID: 19165148
- PMCID: PMC2657586
- DOI: 10.1038/emboj.2009.4
Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis
Abstract
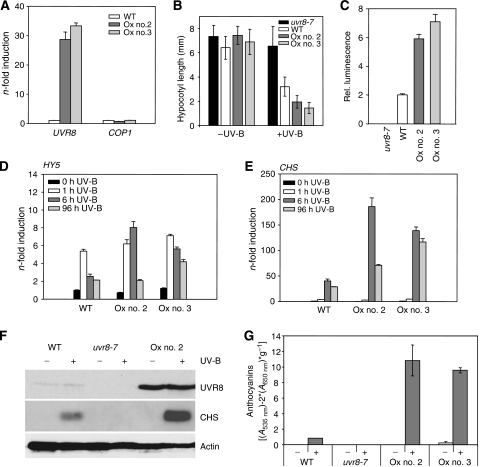
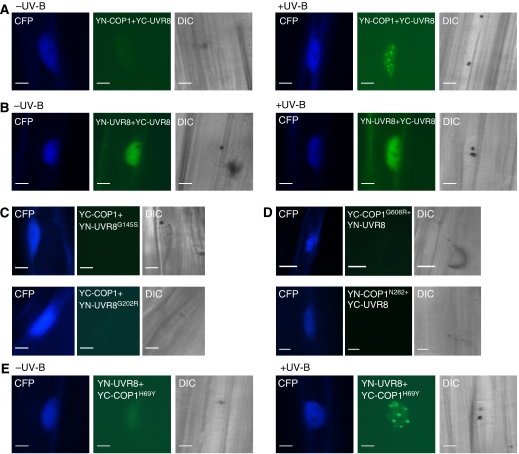
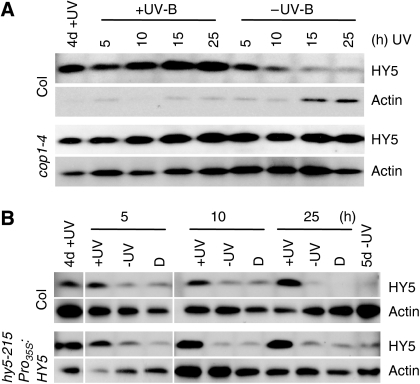
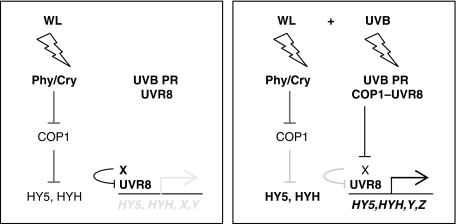
The ultraviolet-B (UV-B) portion of the solar radiation functions as an environmental signal for which plants have evolved specific and sensitive UV-B perception systems. The UV-B-specific UV RESPONSE LOCUS 8 (UVR8) and the multifunctional E3 ubiquitin ligase CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1) are key regulators of the UV-B response. We show here that uvr8-null mutants are deficient in UV-B-induced photomorphogenesis and hypersensitive to UV-B stress, whereas overexpression of UVR8 results in enhanced UV-B photomorphogenesis, acclimation and tolerance to UV-B stress. By using sun simulators, we provide evidence at the physiological level that UV-B acclimation mediated by the UV-B-specific photoregulatory pathway is indeed required for survival in sunlight. At the molecular level, we demonstrate that the wild type but not the mutant UVR8 and COP1 proteins directly interact in a UV-B-dependent, rapid manner in planta. These data collectively suggest that UV-B-specific interaction of COP1 and UVR8 in the nucleus is a very early step in signalling and responsible for the plant's coordinated response to UV-B ensuring UV-B acclimation and protection in the natural environment.
Figures




Similar articles
-
C-terminal region of the UV-B photoreceptor UVR8 initiates signaling through interaction with the COP1 protein.Proc Natl Acad Sci U S A. 2012 Oct 2;109(40):16366-70. doi: 10.1073/pnas.1210898109. Epub 2012 Sep 17. Proc Natl Acad Sci U S A. 2012. PMID: 22988111 Free PMC article.
-
Photoactivated UVR8-COP1 module determines photomorphogenic UV-B signaling output in Arabidopsis.PLoS Genet. 2014 Mar 20;10(3):e1004218. doi: 10.1371/journal.pgen.1004218. eCollection 2014 Mar. PLoS Genet. 2014. PMID: 24651064 Free PMC article.
-
Negative feedback regulation of UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis.Proc Natl Acad Sci U S A. 2010 Nov 16;107(46):20132-7. doi: 10.1073/pnas.0914532107. Epub 2010 Nov 1. Proc Natl Acad Sci U S A. 2010. PMID: 21041653 Free PMC article.
-
Perception and Signaling of Ultraviolet-B Radiation in Plants.Annu Rev Plant Biol. 2021 Jun 17;72:793-822. doi: 10.1146/annurev-arplant-050718-095946. Epub 2021 Feb 26. Annu Rev Plant Biol. 2021. PMID: 33636992 Review.
-
Recent advances in UV-B signalling: interaction of proteins with the UVR8 photoreceptor.J Exp Bot. 2025 Feb 7;76(3):873-881. doi: 10.1093/jxb/erae132. J Exp Bot. 2025. PMID: 38525857 Free PMC article. Review.
Cited by
-
The ROS Wheel: Refining ROS Transcriptional Footprints.Plant Physiol. 2016 Jul;171(3):1720-33. doi: 10.1104/pp.16.00420. Epub 2016 May 31. Plant Physiol. 2016. PMID: 27246095 Free PMC article.
-
The UVR8 UV-B Photoreceptor: Perception, Signaling and Response.Arabidopsis Book. 2013 Jun 11;11:e0164. doi: 10.1199/tab.0164. Print 2013. Arabidopsis Book. 2013. PMID: 23864838 Free PMC article.
-
Role of Ubiquitin-Mediated Degradation System in Plant Biology.Front Plant Sci. 2016 Jun 8;7:806. doi: 10.3389/fpls.2016.00806. eCollection 2016. Front Plant Sci. 2016. PMID: 27375660 Free PMC article. Review.
-
Dimer/monomer status and in vivo function of salt-bridge mutants of the plant UV-B photoreceptor UVR8.Plant J. 2016 Oct;88(1):71-81. doi: 10.1111/tpj.13260. Epub 2016 Sep 9. Plant J. 2016. PMID: 27385642 Free PMC article.
-
UV-B Pre-treatment Alters Phenolics Response to Monilinia fructicola Infection in a Structure-Dependent Way in Peach Skin.Front Plant Sci. 2018 Nov 6;9:1598. doi: 10.3389/fpls.2018.01598. eCollection 2018. Front Plant Sci. 2018. PMID: 30459788 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, Gadrinab C, Heller C, Jeske A, Koesema E, Meyers CC, Parker H, Prednis L, Ansari Y, Choy N, Deen H et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Ang LH, Chattopadhyay S, Wei N, Oyama T, Okada K, Batschauer A, Deng XW (1998) Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol Cell 1: 213–222 - PubMed
-
- Boccalandro HE, Rossi MC, Saijo Y, Deng XW, Casal JJ (2004) Promotion of photomorphogenesis by COP1. Plant Mol Biol 56: 905–915 - PubMed
-
- Brosche M, Strid A (2003) Molecular events following perception of ultraviolet-B radiation by plants. Physiol Plant 117: 1–10
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
