

Matrix alkalinization: a novel mitochondrial signal for sustained pancreatic beta-cell activation
- PMID: 19165153
- PMCID: PMC2646158
- DOI: 10.1038/emboj.2008.302
Matrix alkalinization: a novel mitochondrial signal for sustained pancreatic beta-cell activation
Abstract
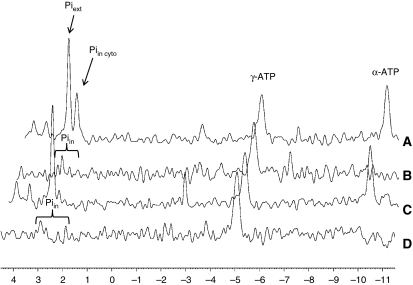
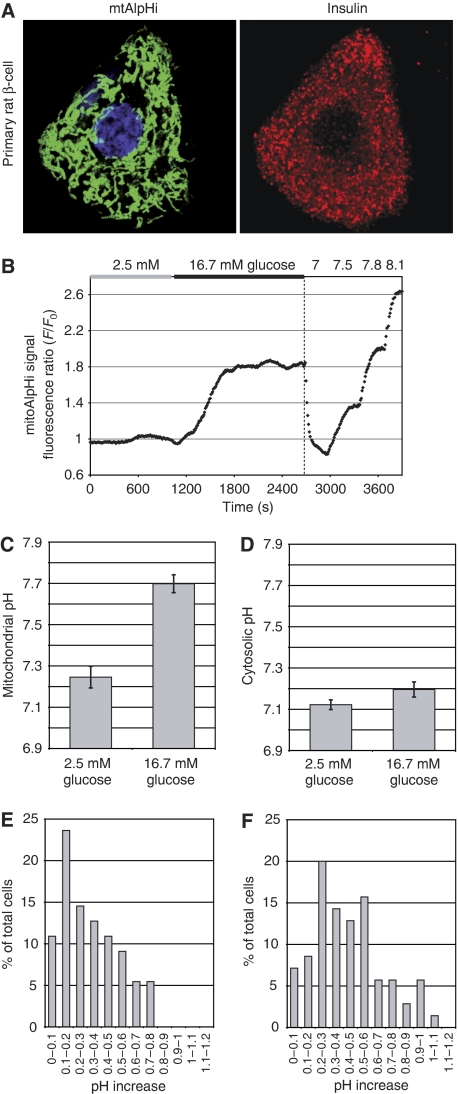
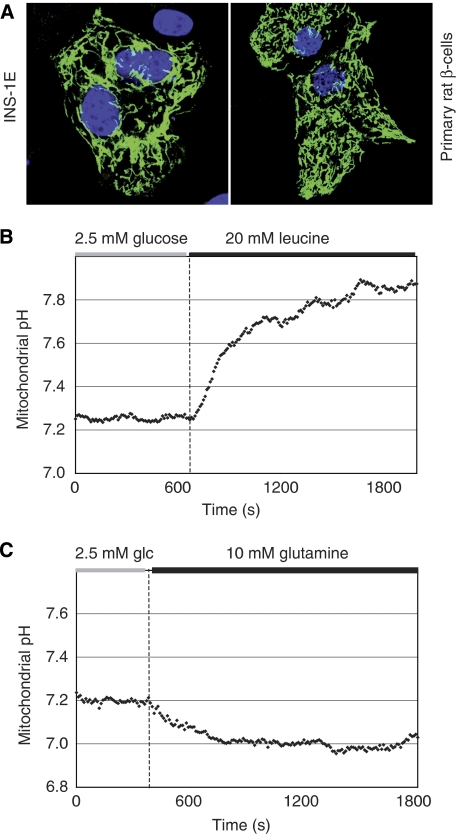
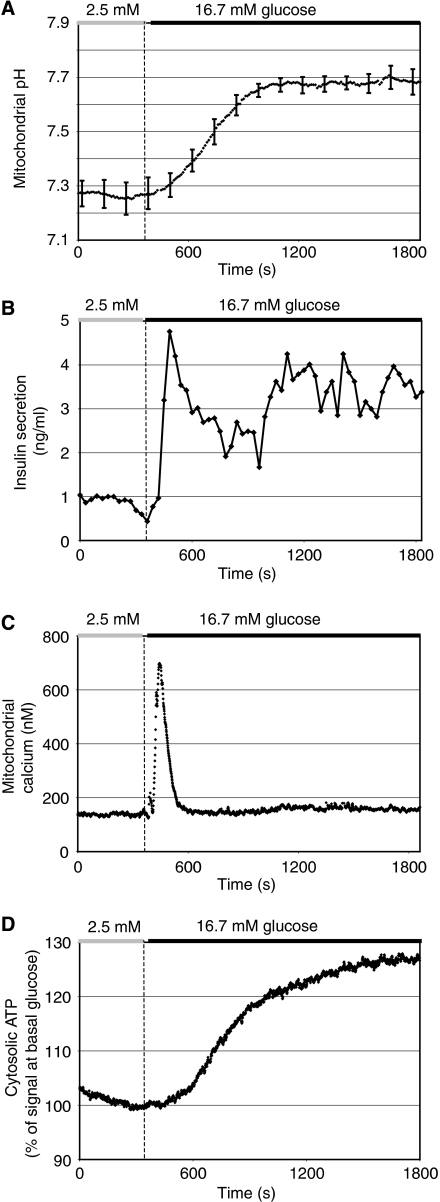
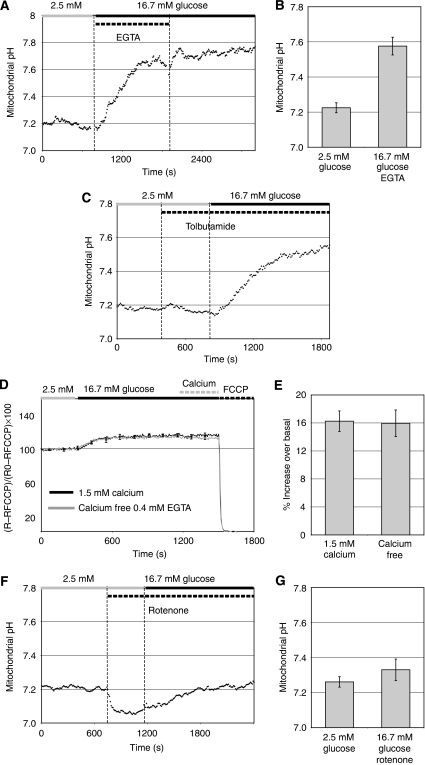
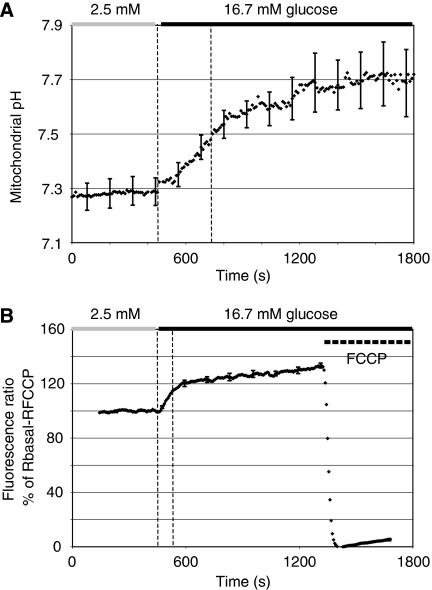
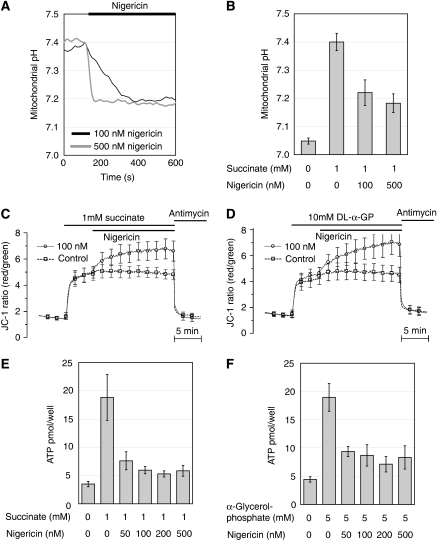
Nutrient secretagogues activate mitochondria of the pancreatic beta-cell through the provision of substrate, hyperpolarisation of the inner mitochondrial membrane and mitochondrial calcium rises. We report that mitochondrial matrix pH, a parameter not previously studied in the beta-cell, also exerts an important control function in mitochondrial metabolism. During nutrient stimulation matrix pH alkalinises, monitored by the mitochondrial targeted fluorescent pH-sensitive protein mtAlpHi or (31)P-NMR inorganic phosphate chemical shifts following saturation transfer. Compared with other cell types, the resting mitochondrial pH was surprisingly low, rising from pH 7.25 to 7.7 during nutrient stimulation of rat beta-cells. As cytosolic alkalinisation to the nutrient was of much smaller amplitude, the matrix alkalinisation was accompanied by a pronounced increase of the DeltapH across the inner mitochondrial membrane. Furthermore, matrix alkalinisation closely correlates with the cytosolic ATP net increase, which is also associated with elevated ATP synthesis rates in mitochondria. Preventing DeltapH increases in permeabilised cells abrogated substrate-driven ATP synthesis. We propose that the mitochondrial pH and DeltapH are key determinants of mitochondrial energy metabolism and metabolite transport important for cell activation.
Figures

Comment in
-
Matrix alkalinisation unleashes β-cell mitochondria.Islets. 2009 Sep-Oct;1(2):154-6. doi: 10.4161/isl.1.2.9058. Islets. 2009. PMID: 21099264
References
-
- Abad MF, Di Benedetto G, Magalhaes PJ, Filippin L, Pozzan T (2004) Mitochondrial pH monitored by a new engineered green fluorescent protein mutant. J Biol Chem 279: 11521–11529 - PubMed
-
- Balut C, vandeVen M, Despa S, Lambrichts I, Ameloot M, Steels P, Smets I (2008) Measurement of cytosolic and mitochondrial pH in living cells during reversible metabolic inhibition. Kidney Int 73: 226–232 - PubMed
-
- Cline GW, Lepine RL, Papas KK, Kibbey RG, Shulman GI (2004) 13C NMR isotopomer analysis of anaplerotic pathways in INS-1 cells. J Biol Chem 279: 44370–44375 - PubMed
-
- Cline GW, Vidal-Puig AJ, Dufour S, Cadman KS, Lowell BB, Shulman GI (2001) In vivo effects of uncoupling protein-3 gene disruption on mitochondrial energy metabolism. J Biol Chem 276: 20240–20244 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
