

A tense situation: forcing tumour progression
- PMID: 19165226
- PMCID: PMC2649117
- DOI: 10.1038/nrc2544
A tense situation: forcing tumour progression
Abstract
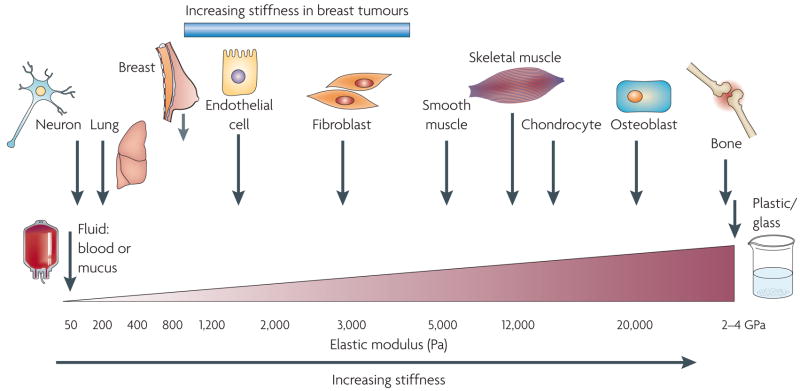
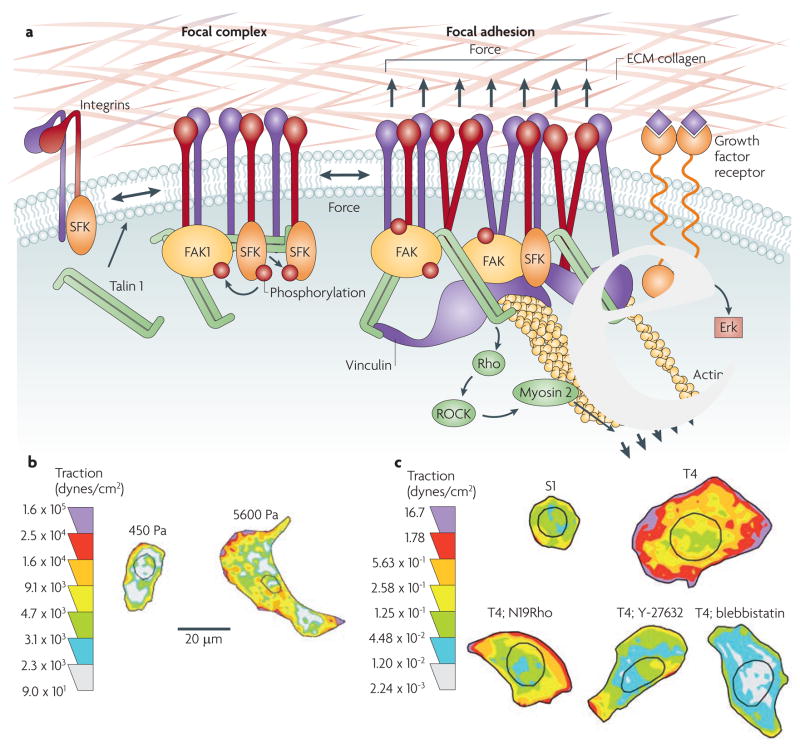
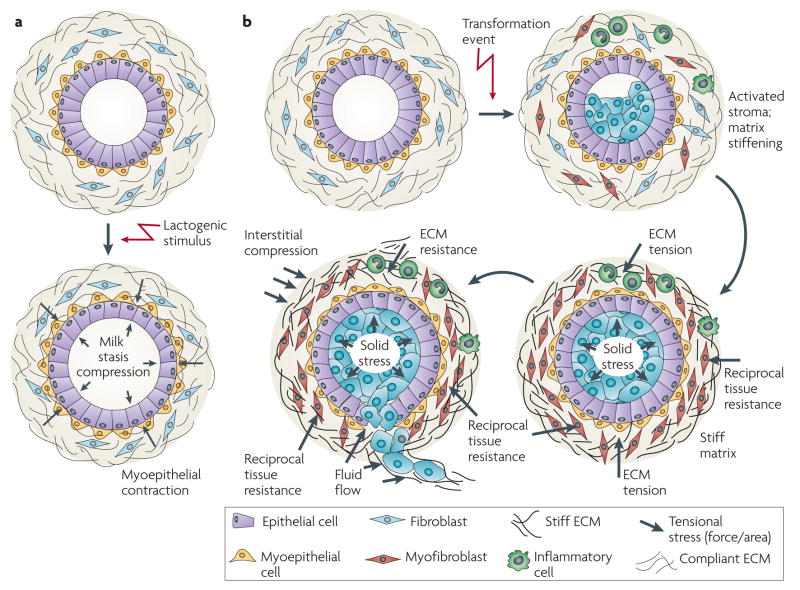
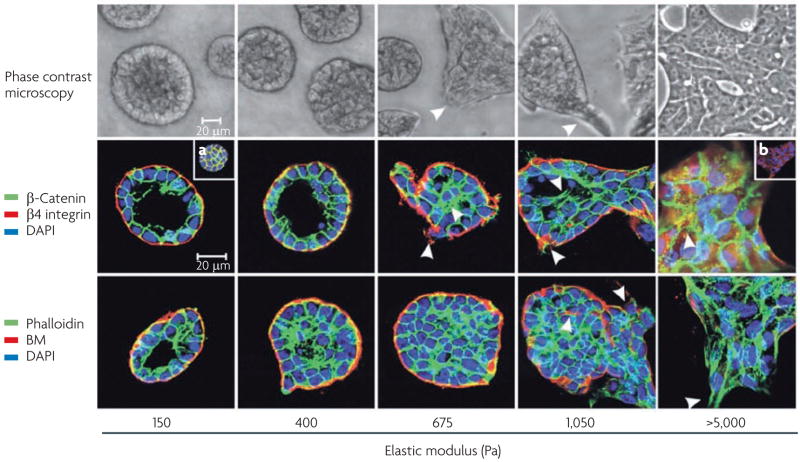
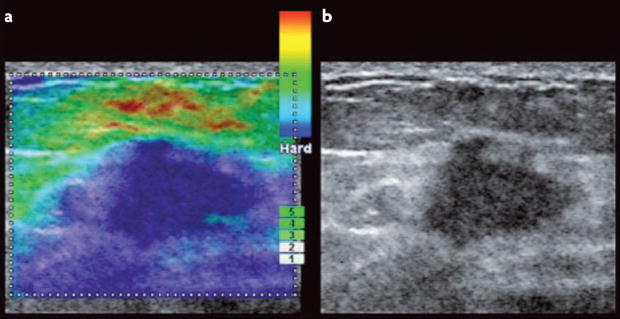
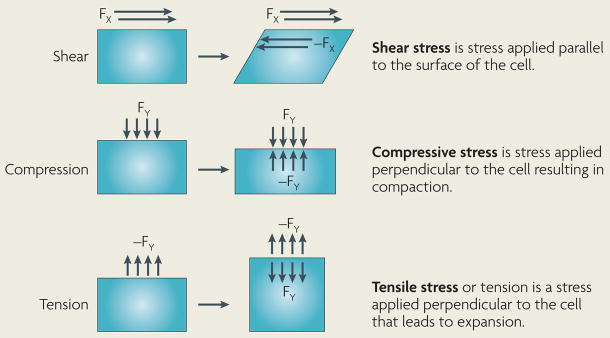
Cells within tissues are continuously exposed to physical forces including hydrostatic pressure, shear stress, and compression and tension forces. Cells dynamically adapt to force by modifying their behaviour and remodelling their microenvironment. They also sense these forces through mechanoreceptors and respond by exerting reciprocal actomyosin- and cytoskeletal-dependent cell-generated force by a process termed 'mechanoreciprocity'. Loss of mechanoreciprocity has been shown to promote the progression of disease, including cancer. Moreover, the mechanical properties of a tissue contribute to disease progression, compromise treatment and might also alter cancer risk. Thus, the changing force that cells experience needs to be considered when trying to understand the complex nature of tumorigenesis.
Figures
References
-
- Gieni RS, Hendzel MJ. Mechanotransduction from the ECM to the genome: are the pieces now in place? J Cell Biochem. 2008;104:1964–1987. - PubMed
-
- Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126:677–689. Contractile myocytes were used to demonstrate that cells sense their mechanical environment. Myotubes form independently of matrix stiffness but myosin–actin striations emerge only on gels with stiffness typical of normal muscle, and not on matrices that are softer or stiffer. - PubMed
-
- McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS. Cell shape, cytoskeletal tension, and rhoa regulate stem cell lineage commitment. Dev Cell. 2004;6:483–495. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
