

Nuclear envelope remnants: fluid membranes enriched in sterols and polyphosphoinositides
- PMID: 19165341
- PMCID: PMC2626249
- DOI: 10.1371/journal.pone.0004255
Nuclear envelope remnants: fluid membranes enriched in sterols and polyphosphoinositides
Abstract
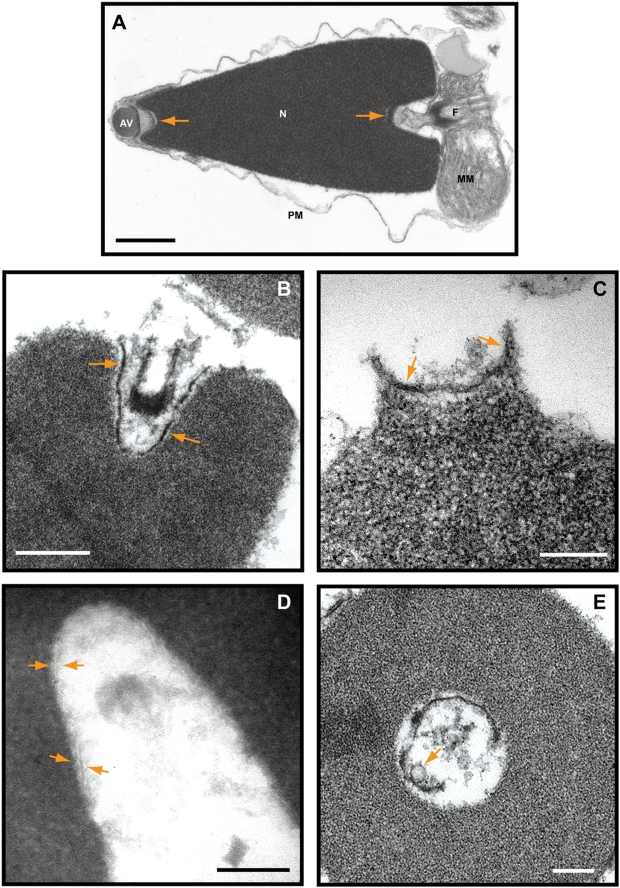
Background: The cytoplasm of eukaryotic cells is a highly dynamic compartment where membranes readily undergo fission and fusion to reorganize the cytoplasmic architecture, and to import, export and transport various cargos within the cell. The double membrane of the nuclear envelope that surrounds the nucleus, segregates the chromosomes from cytoplasm and regulates nucleocytoplasmic transport through pores. Many details of its formation are still unclear. At fertilization the sperm devoid of nuclear envelope pores enters the egg. Although most of the sperm nuclear envelope disassembles, remnants of the envelope at the acrosomal and centriolar fossae do not and are subsequently incorporated into the newly forming male pronuclear envelope. Remnants are conserved from annelid to mammalian sperm.
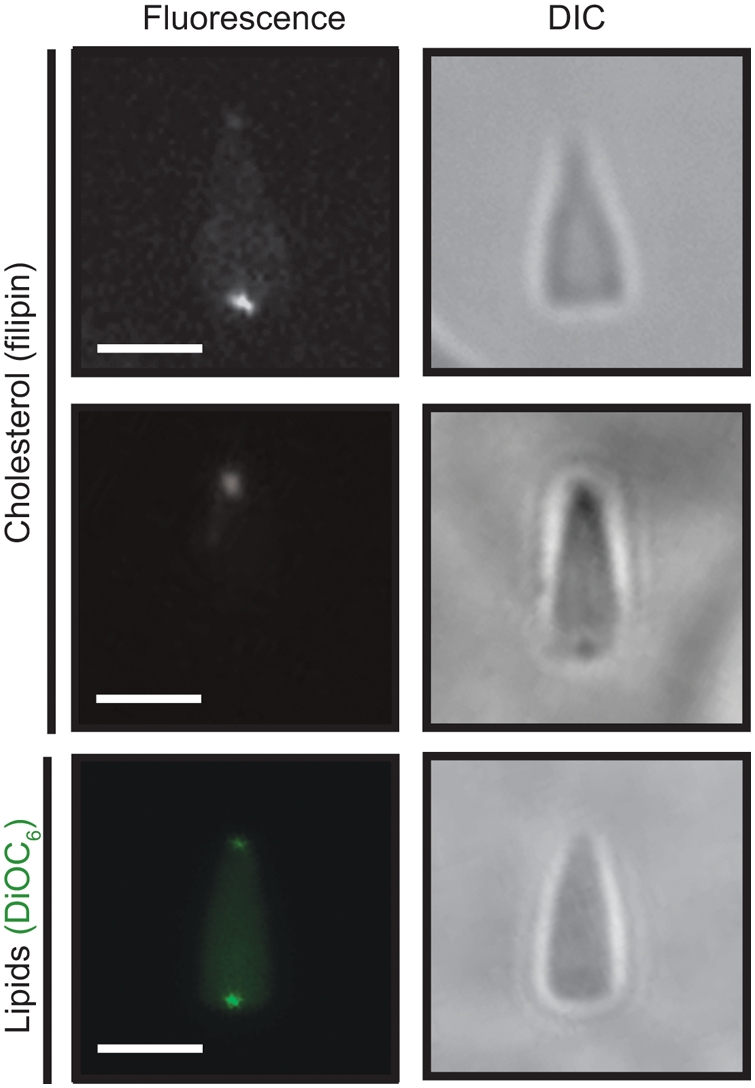
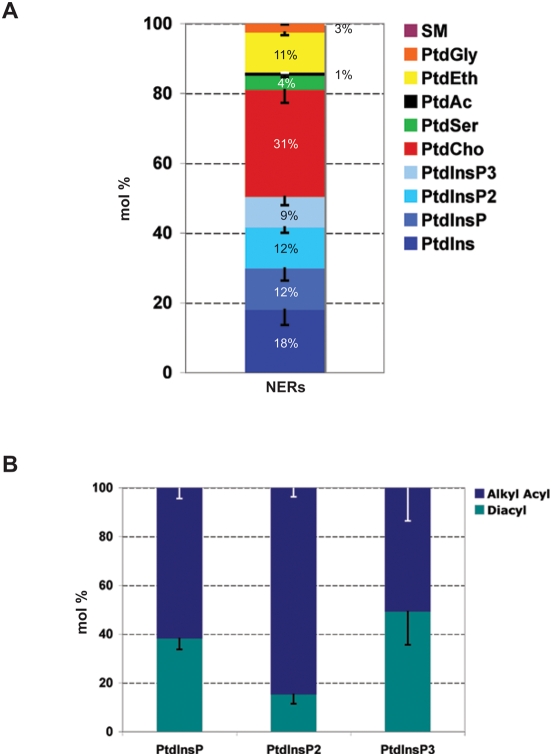
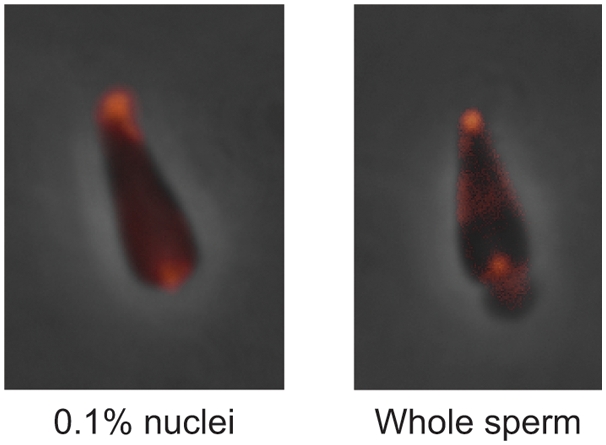
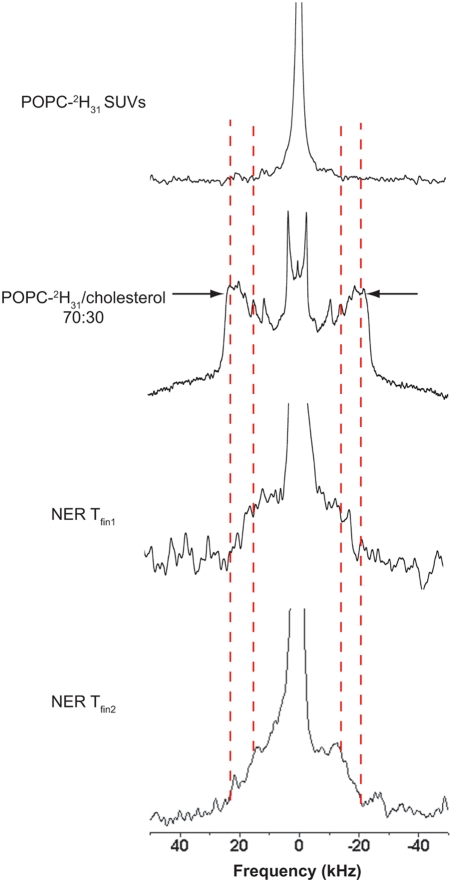
Methodology/principal findings: Using lipid mass spectrometry and a new application of deuterium solid-state NMR spectroscopy we have characterized the lipid composition and membrane dynamics of the sperm nuclear envelope remnants in isolated sperm nuclei.
Conclusions/significance: We report nuclear envelope remnants are relatively fluid membranes rich in sterols, devoid of sphingomyelin, and highly enriched in polyphosphoinositides and polyunsaturated phospholipids. The localization of the polybasic effector domain of MARCKS illustrates the non-nuclear aspect of the polyphosphoinositides. Based on their atypical biophysical characteristics and phospholipid composition, we suggest a possible role for nuclear envelope remnants in membrane fusion leading to nuclear envelope assembly.
Conflict of interest statement
Figures
References
-
- Collas P, Poccia DL. Conserved binding recognition elements of sperm chromatin, sperm lipophilic structures and nuclear envelope precursor vesicles. Eur J Cell Biol. 1996;71:22–32. - PubMed
-
- Collas P, Poccia D. Lipophilic organizing structures of sperm nuclei target membrane vesicle binding and are incorporated into the nuclear envelope. Dev Biol. 1995;169:123–135. - PubMed
-
- Cameron LA, Poccia DL. In vitro development of the sea urchin male pronucleus. Dev Biol. 1994;162:568–578. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
