

A novel function for fragile X mental retardation protein in translational activation
- PMID: 19166269
- PMCID: PMC2628407
- DOI: 10.1371/journal.pbio.1000016
A novel function for fragile X mental retardation protein in translational activation
Abstract
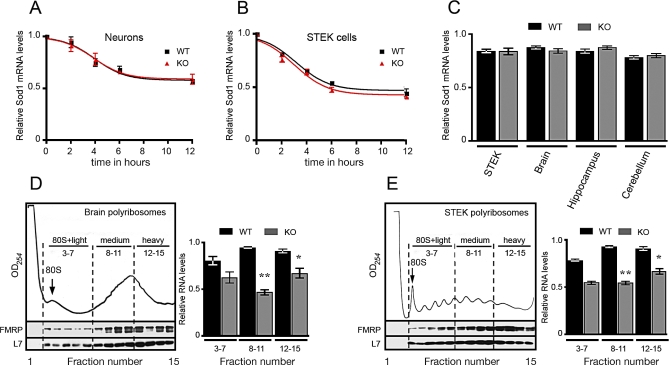
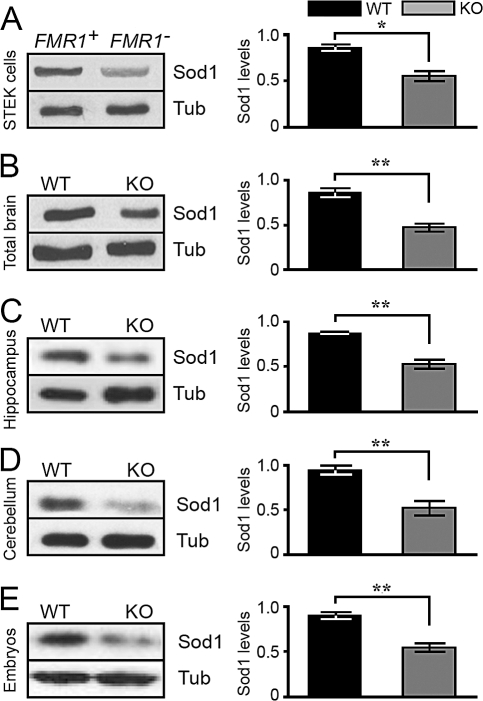
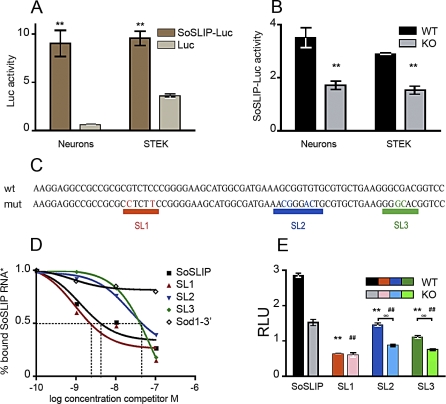
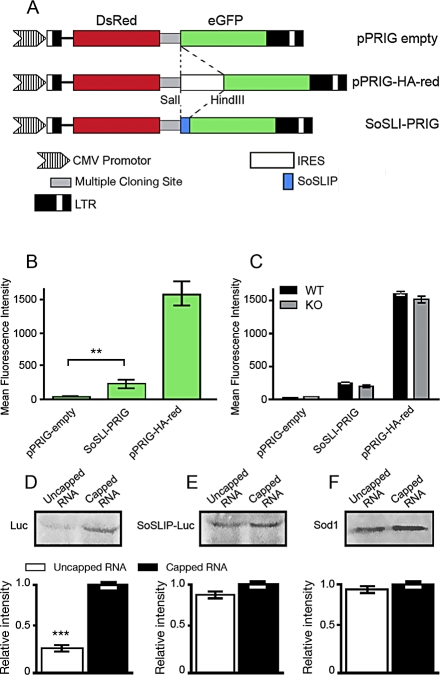
Fragile X syndrome, the most frequent form of inherited mental retardation, is due to the absence of Fragile X Mental Retardation Protein (FMRP), an RNA-binding protein involved in several steps of RNA metabolism. To date, two RNA motifs have been found to mediate FMRP/RNA interaction, the G-quartet and the "kissing complex," which both induce translational repression in the presence of FMRP. We show here a new role for FMRP as a positive modulator of translation. FMRP specifically binds Superoxide Dismutase 1 (Sod1) mRNA with high affinity through a novel RNA motif, SoSLIP (Sod1 mRNA Stem Loops Interacting with FMRP), which is folded as three independent stem-loop structures. FMRP induces a structural modification of the SoSLIP motif upon its interaction with it. SoSLIP also behaves as a translational activator whose action is potentiated by the interaction with FMRP. The absence of FMRP results in decreased expression of Sod1. Because it has been observed that brain metabolism of FMR1 null mice is more sensitive to oxidative stress, we propose that the deregulation of Sod1 expression may be at the basis of several traits of the physiopathology of the Fragile X syndrome, such as anxiety, sleep troubles, and autism.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
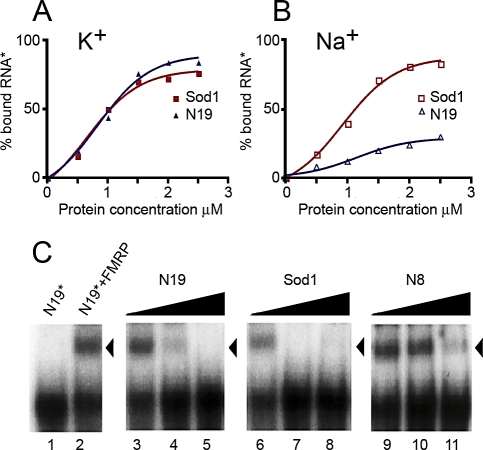
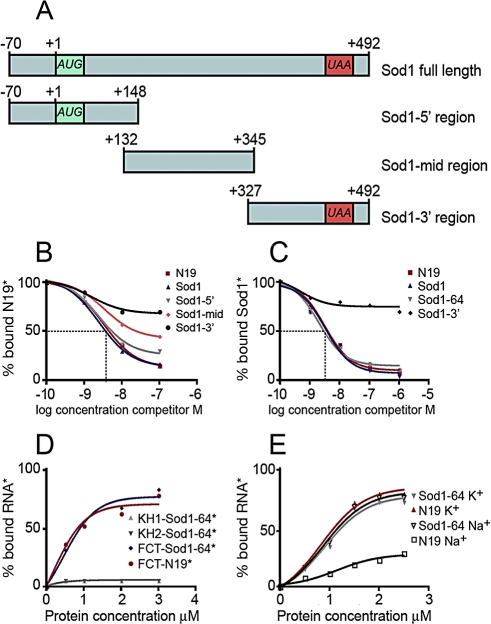
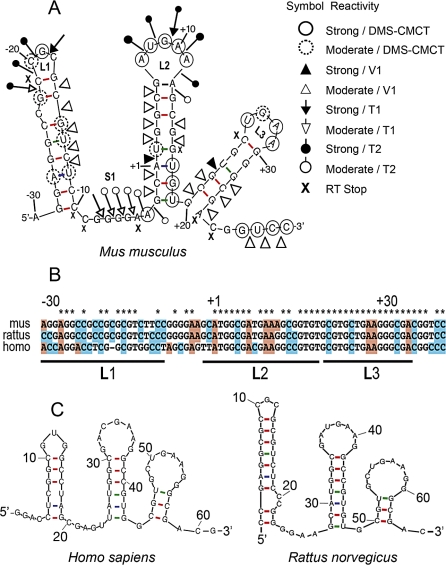
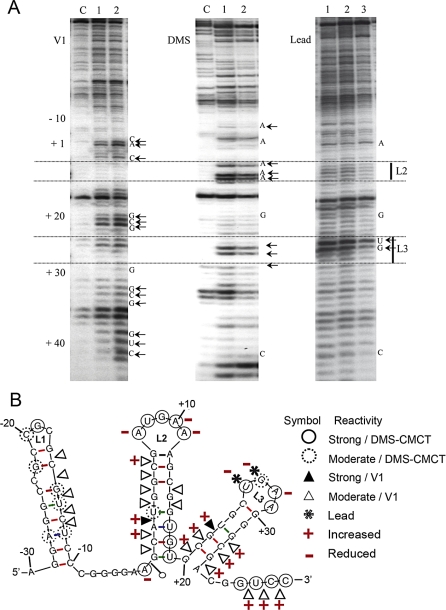
References
-
- Bardoni B, Davidovic L, Bensaid M, Khandjian EW. The fragile X syndrome: exploring its molecular basis and seeking a treatment. Expert Rev Mol Med. 2006;8:1–16. - PubMed
-
- Darnell JC, Jensen KB, Jin P, Brown V, Warren ST, et al. Fragile X mental retardation protein targets G-quartet mRNAs important for neuronal function. Cell. 2001;107:489–499. - PubMed
-
- Castets M, Schaeffer C, Bechara E, Schenck A, Khandjian EW, et al. FMRP interferes with Rac1 pathway and controls actin cytoskeleton dynamics in murine fibroblasts. Hum Mol Genet. 2005;14:835–844. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
