

Glutathione dysregulation and the etiology and progression of human diseases
- PMID: 19166318
- PMCID: PMC2756154
- DOI: 10.1515/BC.2009.033
Glutathione dysregulation and the etiology and progression of human diseases
Abstract
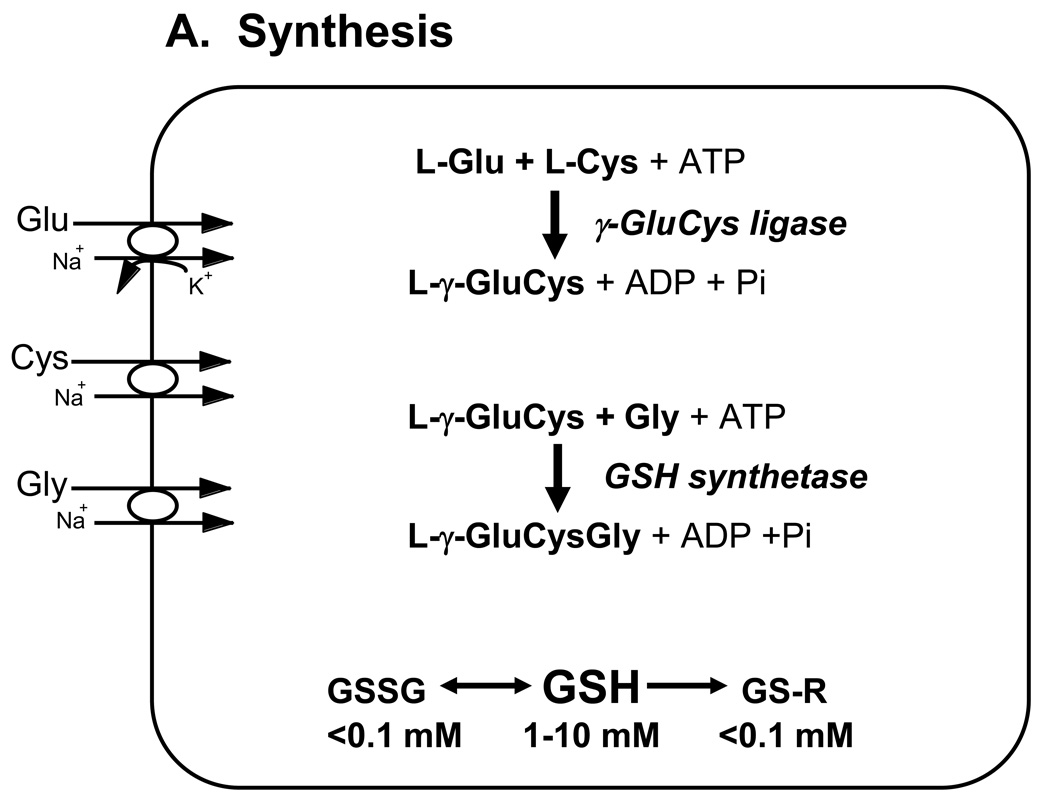
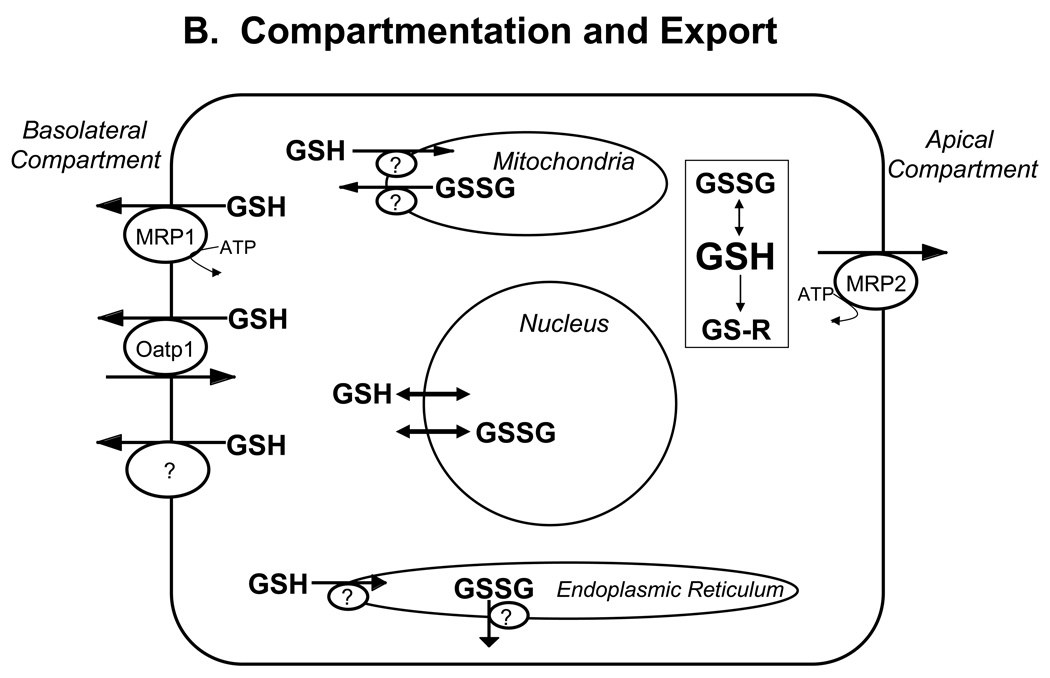
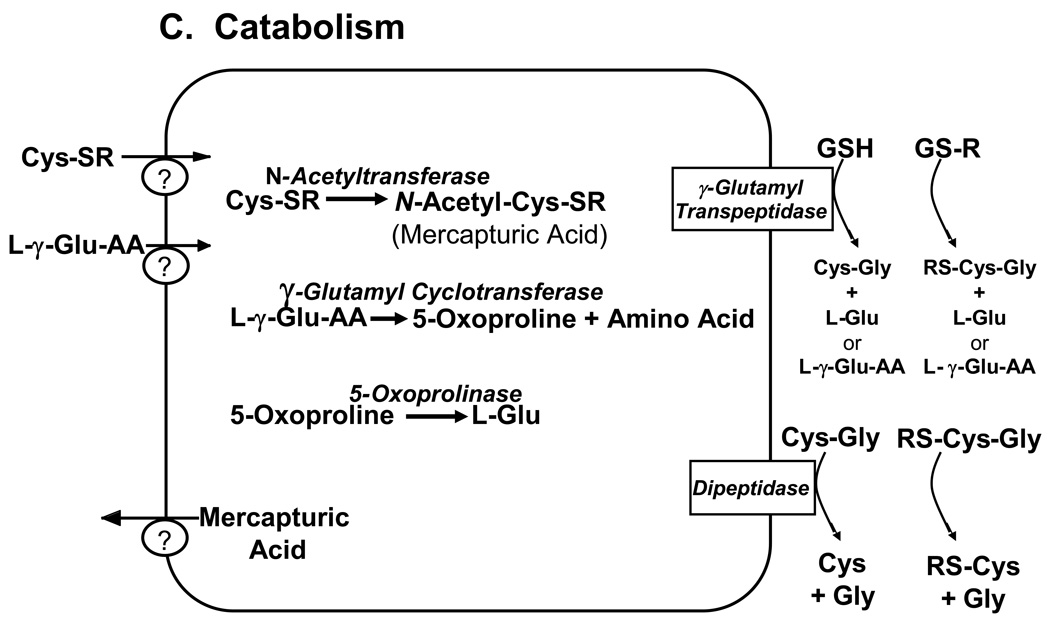
Glutathione (GSH) plays an important role in a multitude of cellular processes, including cell differentiation, proliferation, and apoptosis, and as a result, disturbances in GSH homeostasis are implicated in the etiology and/or progression of a number of human diseases, including cancer, diseases of aging, cystic fibrosis, and cardiovascular, inflammatory, immune, metabolic, and neurodegenerative diseases. Owing to the pleiotropic effects of GSH on cell functions, it has been quite difficult to define the role of GSH in the onset and/or the expression of human diseases, although significant progress is being made. GSH levels, turnover rates, and/or oxidation state can be compromised by inherited or acquired defects in the enzymes, transporters, signaling molecules, or transcription factors that are involved in its homeostasis, or from exposure to reactive chemicals or metabolic intermediates. GSH deficiency or a decrease in the GSH/glutathione disulfide ratio manifests itself largely through an increased susceptibility to oxidative stress, and the resulting damage is thought to be involved in diseases, such as cancer, Parkinson's disease, and Alzheimer's disease. In addition, imbalances in GSH levels affect immune system function, and are thought to play a role in the aging process. Just as low intracellular GSH levels decrease cellular antioxidant capacity, elevated GSH levels generally increase antioxidant capacity and resistance to oxidative stress, and this is observed in many cancer cells. The higher GSH levels in some tumor cells are also typically associated with higher levels of GSH-related enzymes and transporters. Although neither the mechanism nor the implications of these changes are well defined, the high GSH content makes cancer cells chemoresistant, which is a major factor that limits drug treatment. The present report highlights and integrates the growing connections between imbalances in GSH homeostasis and a multitude of human diseases.
Figures

References
-
- Adams JD, Jr, Klaidman LK, Odunze IN, Shen HC, Miller CA. Alzheimer's and Parkinson's disease. Brain levels of glutathione, glutathione disulfide, and vitamin E. Mol. Chem. Neuropathol. 1991;14:213–226. - PubMed
-
- Akahoshi M, Nakashima H, Tanaka Y, Kohsaka T, Nagano S, Ohgami E, Arinobu Y, Yamaoka K, Niiro H, Shinozaki M, Hirakata H, Horiuchi T, Otsuka T, Niho Y. Th1/Th2 balance of peripheral T helper cells in systemic lupus erythematosus. Arthritis. Rheum. 1999;42:1644–1648. - PubMed
-
- Andersson C, Gaston B, Roomans GM. S-Nitrosoglutathione induces functional DeltaF508-CFTR in airway epithelial cells. Biochem Biophys Res Commun. 2002;297:552–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK048823/DK/NIDDK NIH HHS/United States
- ES03828/ES/NIEHS NIH HHS/United States
- R21 ES017470/ES/NIEHS NIH HHS/United States
- ES01247/ES/NIEHS NIH HHS/United States
- P30 ES001247/ES/NIEHS NIH HHS/United States
- P30 ES003828/ES/NIEHS NIH HHS/United States
- ES07026/ES/NIEHS NIH HHS/United States
- R01 ES007026/ES/NIEHS NIH HHS/United States
- R25 ES016254/ES/NIEHS NIH HHS/United States
- R01 ES014899/ES/NIEHS NIH HHS/United States
- ES16254/ES/NIEHS NIH HHS/United States
- DK48823/DK/NIDDK NIH HHS/United States
- T32 ES007026/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources