

Regulation of cell cytoskeleton and membrane mechanics by electric field: role of linker proteins
- PMID: 19167316
- PMCID: PMC2716454
- DOI: 10.1016/j.bpj.2008.09.035
Regulation of cell cytoskeleton and membrane mechanics by electric field: role of linker proteins
Abstract
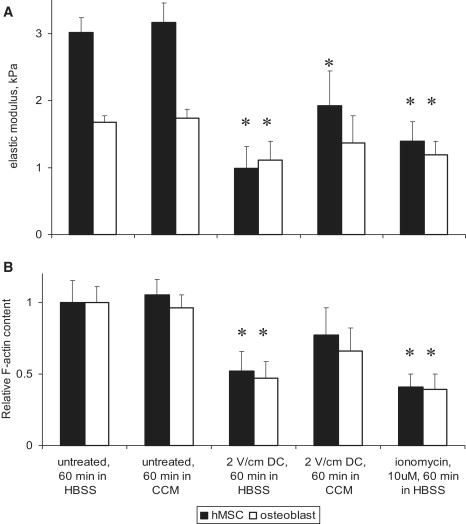
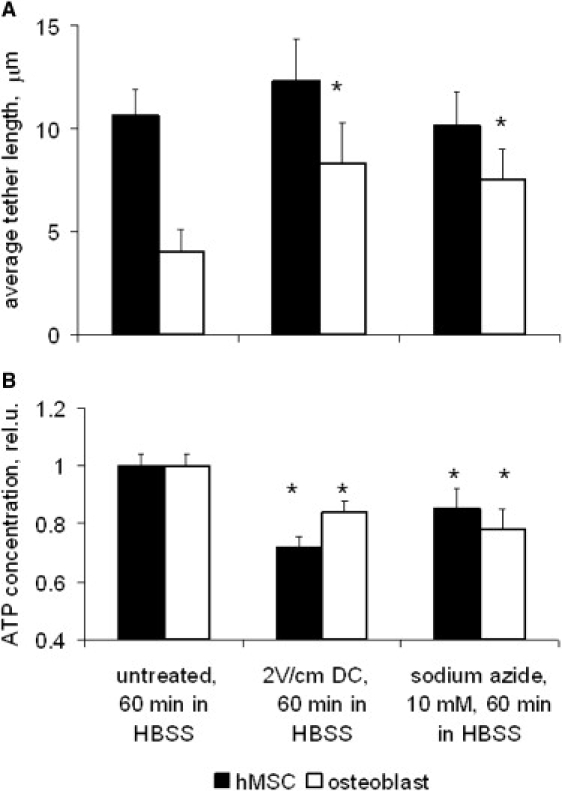
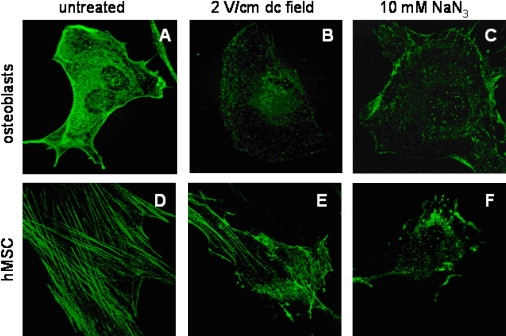
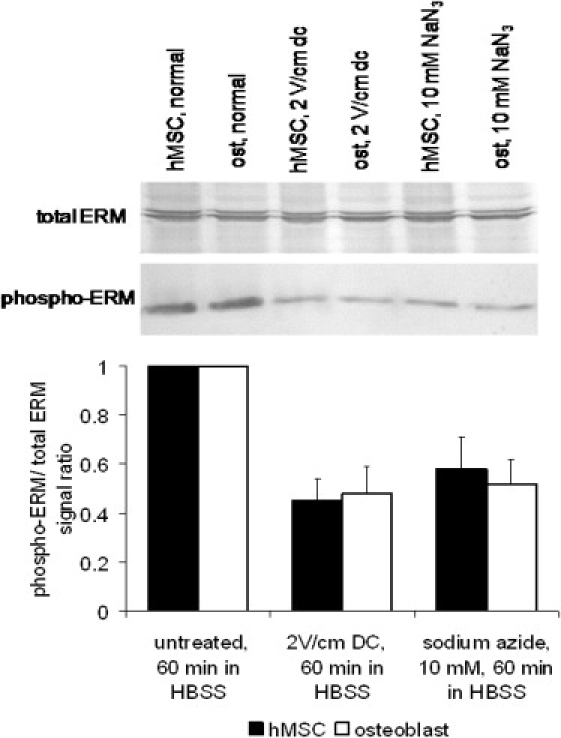
Cellular mechanics is known to play an important role in the cell homeostasis including proliferation, motility, and differentiation. Significant variation in the mechanical properties between different cell types suggests that control of the cell metabolism is feasible through manipulation of the cell mechanical parameters using external physical stimuli. We investigated the electrocoupling mechanisms of cellular biomechanics modulation by an electrical stimulation in two mechanically distinct cell types--human mesenchymal stem cells and osteoblasts. Application of a 2 V/cm direct current electric field resulted in approximately a twofold decrease in the cell elasticity and depleted intracellular ATP. Reduction in the ATP level led to inhibition of the linker proteins that are known to physically couple the cell membrane and cytoskeleton. The membrane separation from the cytoskeleton was confirmed by up to a twofold increase in the membrane tether length that was extracted from the cell membrane after an electrical stimulation. In comparison to human mesenchymal stem cells, the membrane-cytoskeleton attachment in osteoblasts was much stronger but, in response to the same electrical stimulation, the membrane detachment from the cytoskeleton was found to be more pronounced. The observed effects mediated by an electric field are cell type- and serum-dependent and can potentially be used for electrically assisted cell manipulation. An in-depth understanding and control of the mechanisms to regulate cell mechanics by external physical stimulus (e.g., electric field) may have great implications for stem cell-based tissue engineering and regenerative medicine.
Figures


References
-
- Boscolo P., Di Gioacchino M., Di Giampaolo L., Antonucci A., Di Luzio S. Combined effects of electromagnetic fields on immune and nervous responses. Int. J. Immunopathol. Pharmacol. 2007;20:59–63. - PubMed
-
- Mycielska M.E., Djamgoz M.B. Cellular mechanisms of direct-current electric field effects: galvanotaxis and metastatic disease. J. Cell Sci. 2004;117:1631–1639. - PubMed
-
- Song B., Zhao M., Forrester J., McCaig C. Nerve regeneration and wound healing are stimulated and directed by an endogenous electrical field in vivo. J. Cell Sci. 2004;117:4681–4690. - PubMed
-
- Hotary K.B., Robinson K.R. Endogenous electrical currents and voltage gradients in Xenopus embryos and the consequences of their disruption. Dev. Biol. 1994;166:789–800. - PubMed
-
- Revest P.A., Jones H.C., Abbott N.J. Transendothelial electrical potential across pial vessels in anesthetized rats: a study of ion permeability and transport at the blood-brain barrier. Brain Res. 1994;652:76–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
