

Digital signaling and hysteresis characterize ras activation in lymphoid cells
- PMID: 19167334
- PMCID: PMC2662698
- DOI: 10.1016/j.cell.2008.11.051
Digital signaling and hysteresis characterize ras activation in lymphoid cells
Abstract
Activation of Ras proteins underlies functional decisions in diverse cell types. Two molecules, RasGRP and SOS, catalyze Ras activation in lymphocytes. Binding of active Ras to SOS' allosteric pocket markedly increases SOS' activity establishing a positive feedback loop for SOS-mediated Ras activation. Integrating in silico and in vitro studies, we demonstrate that digital signaling in lymphocytes (cells are "on" or "off") is predicated upon feedback regulation of SOS. SOS' feedback loop leads to hysteresis in the dose-response curve, which can enable a capacity to sustain Ras activation as stimuli are withdrawn and exhibit "memory" of past encounters with antigen. Ras activation via RasGRP alone is analog (graded increase in amplitude with stimulus). We describe how complementary analog (RasGRP) and digital (SOS) pathways act on Ras to efficiently convert analog input to digital output. Numerous predictions regarding the impact of our findings on lymphocyte function and development are noted.
Figures
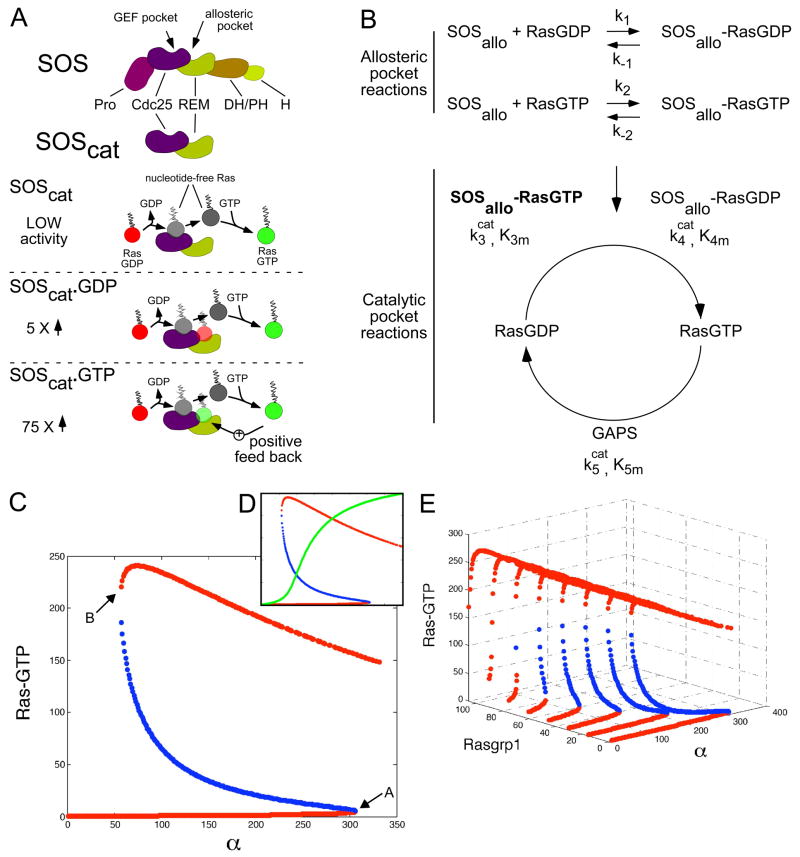
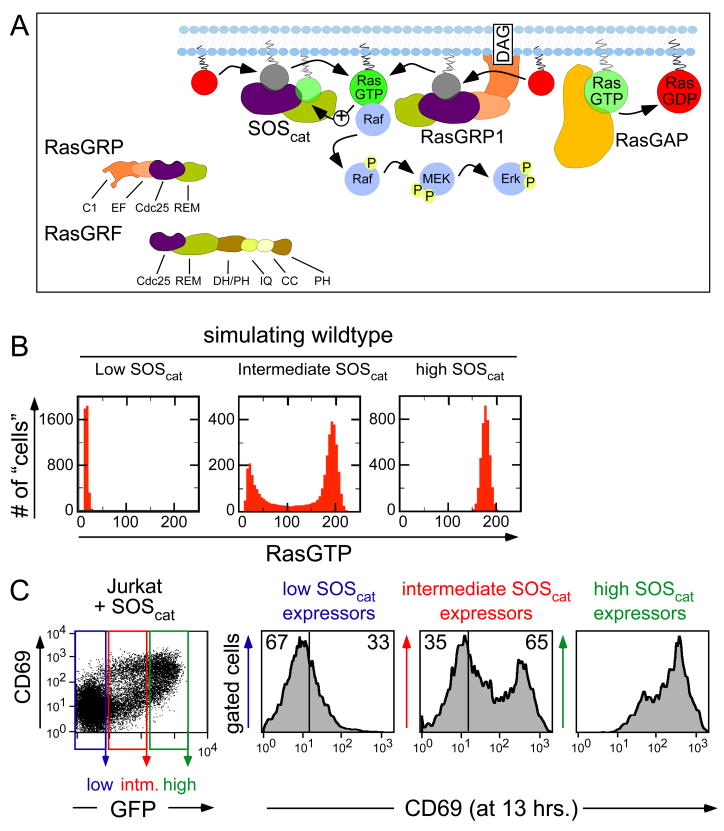
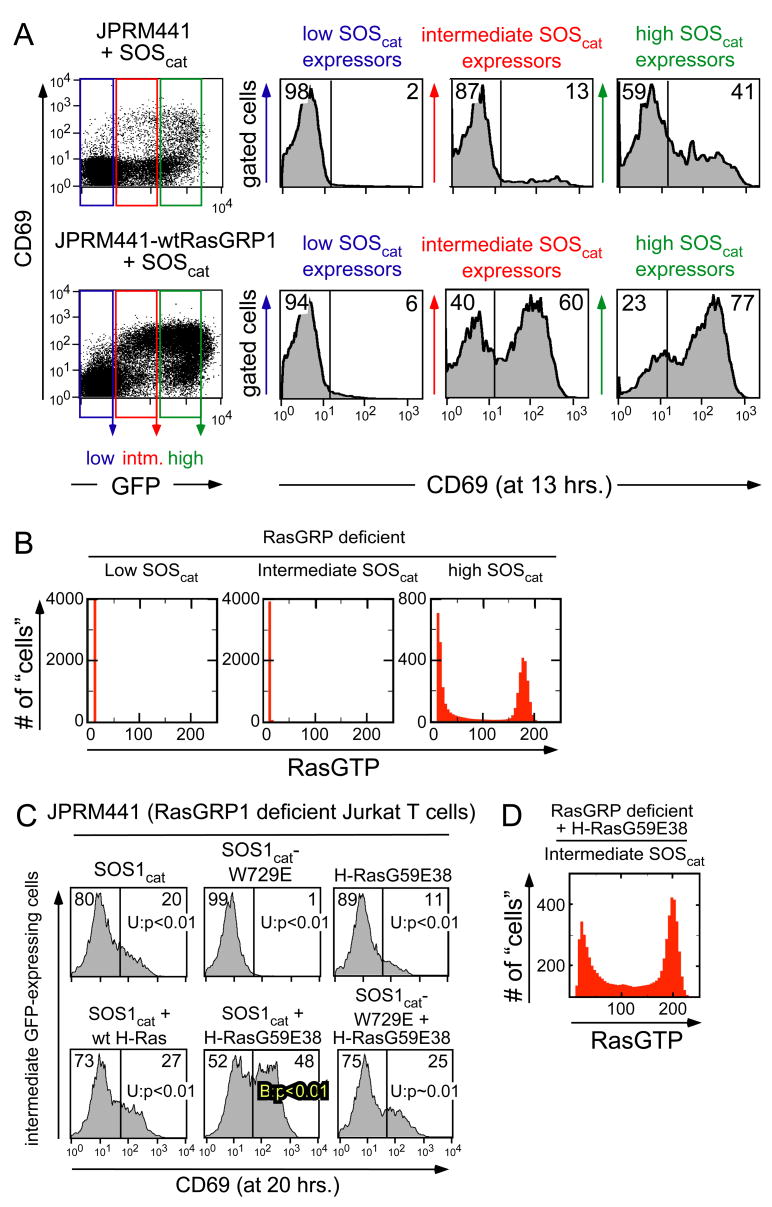
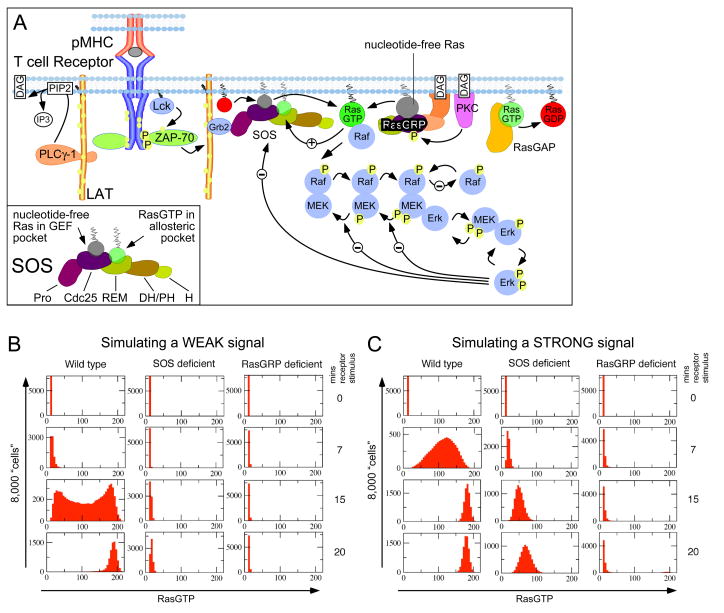
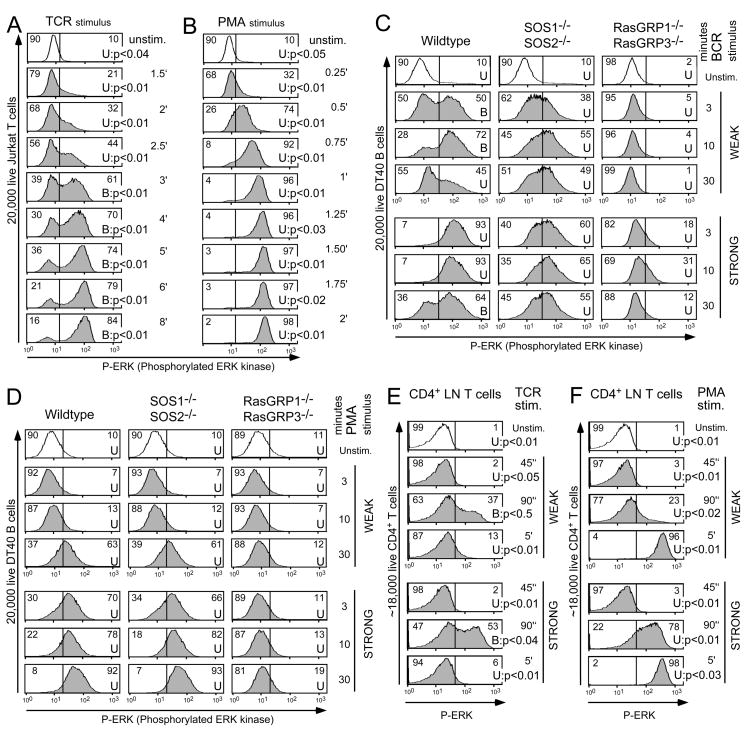
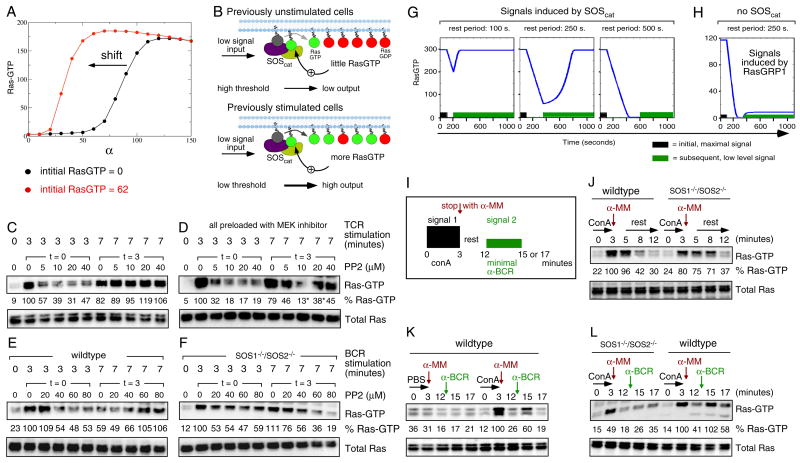
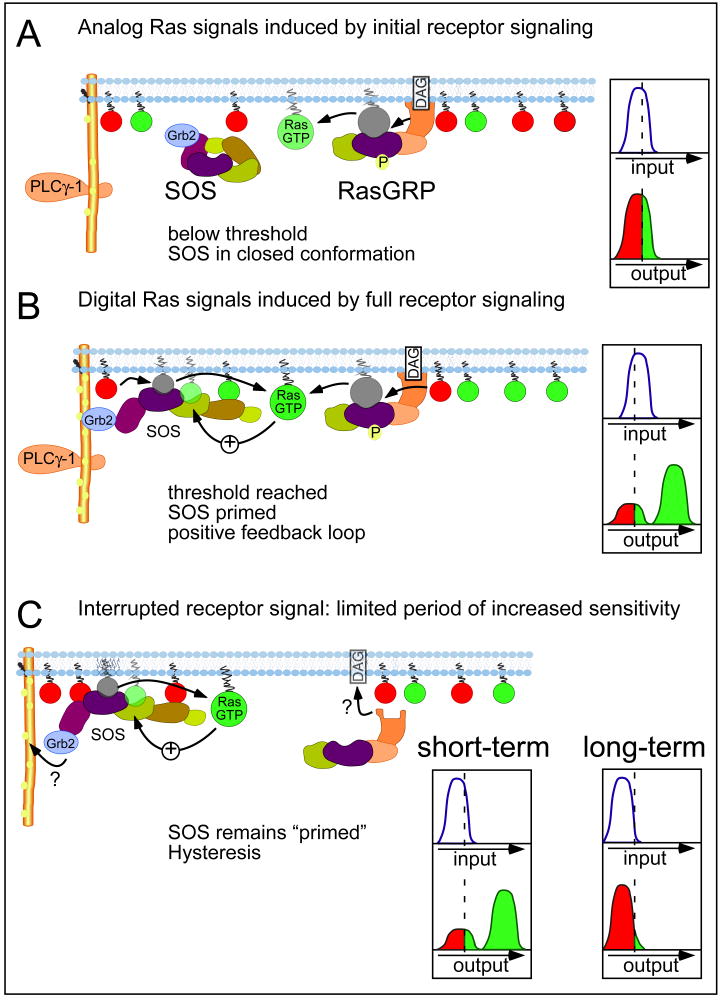
References
-
- Barkai N, Leibler S. Circadian clocks limited by noise. Nature. 2000;403:267–268. - PubMed
-
- Bhalla US, Iyengar R. Emergent properties of networks of biological signaling pathways. Science. 1999;283:381–387. - PubMed
-
- Bortz BKMH, Lebowitz JL. Journal of Computational Physics. 1975;17
-
- Bousso P, Robey E. Dynamics of CD8(+) T cell priming by dendritic cells in intact lymph nodes. Nature immunology. 2003;4:579–585. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
