

Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake
- PMID: 19168643
- PMCID: PMC2649387
- DOI: 10.1104/pp.108.133348
Nitric oxide contributes to cadmium toxicity in Arabidopsis by promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake
Abstract
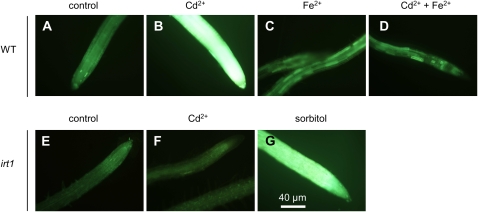
Nitric oxide (NO) functions as a cell-signaling molecule in plants. In particular, a role for NO in the regulation of iron homeostasis and in the plant response to toxic metals has been proposed. Here, we investigated the synthesis and the role of NO in plants exposed to cadmium (Cd(2+)), a nonessential and toxic metal. We demonstrate that Cd(2+) induces NO synthesis in roots and leaves of Arabidopsis (Arabidopsis thaliana) seedlings. This production, which is sensitive to NO synthase inhibitors, does not involve nitrate reductase and AtNOA1 but requires IRT1, encoding a major plasma membrane transporter for iron but also Cd(2+). By analyzing the incidence of NO scavenging or inhibition of its synthesis during Cd(2+) treatment, we demonstrated that NO contributes to Cd(2+)-triggered inhibition of root growth. To understand the mechanisms underlying this process, a microarray analysis was performed in order to identify NO-modulated root genes up- and down-regulated during Cd(2+) treatment. Forty-three genes were identified encoding proteins related to iron homeostasis, proteolysis, nitrogen assimilation/metabolism, and root growth. These genes include IRT1. Investigation of the metal and ion contents in Cd(2+)-treated roots in which NO synthesis was impaired indicates that IRT1 up-regulation by NO was consistently correlated to NO's ability to promote Cd(2+) accumulation in roots. This analysis also highlights that NO is responsible for Cd(2+)-induced inhibition of root Ca(2+) accumulation. Taken together, our results suggest that NO contributes to Cd(2+) toxicity by favoring Cd(2+) versus Ca(2+) uptake and by initiating a cellular pathway resembling those activated upon iron deprivation.
Figures







Comment in
-
NO contributes to cadmium toxicity in Arabidopsis thaliana by mediating an iron deprivation response.Plant Signal Behav. 2009 Mar;4(3):252-4. doi: 10.4161/psb.4.3.8032. Plant Signal Behav. 2009. PMID: 19721766 Free PMC article.
References
-
- Arnaud N, Murgia I, Boucherez J, Briat JF, Cellier F, Gaymard F (2006) An iron-induced nitric oxide burst precedes ubiquitin-dependent protein degradation for Arabidopsis AtFer1 ferritin gene expression. J Biol Chem 281 23579–23588 - PubMed
-
- Badri DV, Loyola-Vargas VM, Du J, Stermitz FR, Broeckling CD, Iglesias-Andreu L, Vivanco JM (2008) Transcriptome analysis of Arabidopsis roots treated with signaling compounds: a focus on signal transduction, metabolic regulation and secretion. New Phytol 179 209–223 - PubMed
-
- Bartha B, Kolbert Z, Erdei L (2005) Nitric oxide production induced by heavy metals in Brassica juncea L. Czern. and Pisum sativum L. Acta Biol Szegediensis 49 9–12
-
- Becher M, Talke IN, Krall L, Kramer U (2004) Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyperaccumulator Arabidopsis halleri. Plant J 37 251–268 - PubMed
-
- Bereczky Z, Wang HY, Schubert V, Ganal M, Bauer P (2003) Differential regulation of nramp and irt metal transporter genes in wild type and iron uptake mutants of tomato. J Biol Chem 278 24697–24704 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
