

Inhibition of aurora kinases for tailored risk-adapted treatment of multiple myeloma
- PMID: 19171872
- PMCID: PMC2700334
- DOI: 10.1182/blood-2008-09-178350
Inhibition of aurora kinases for tailored risk-adapted treatment of multiple myeloma
Abstract
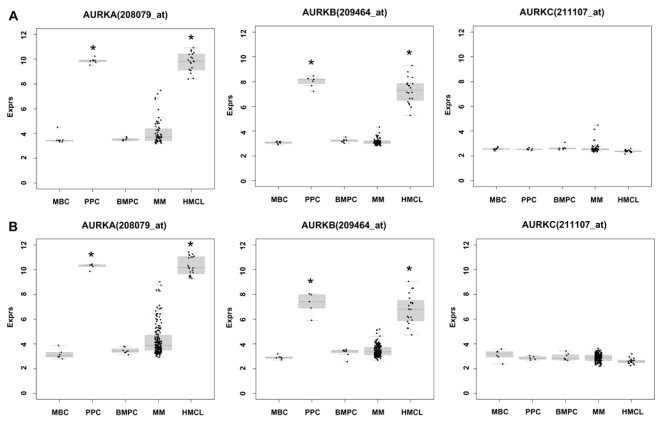
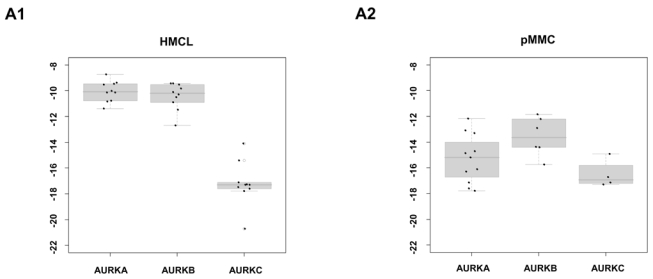
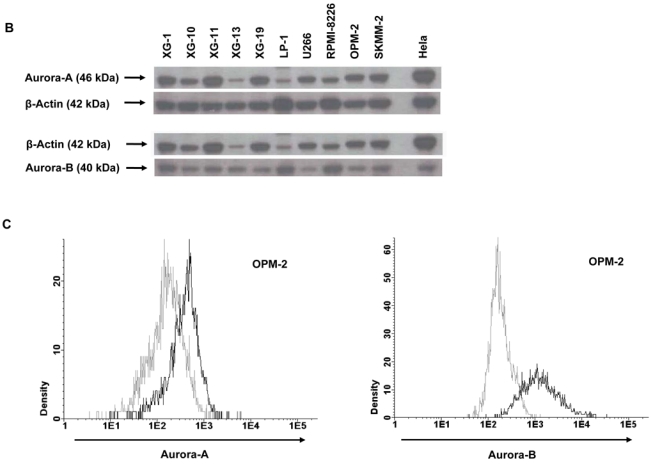
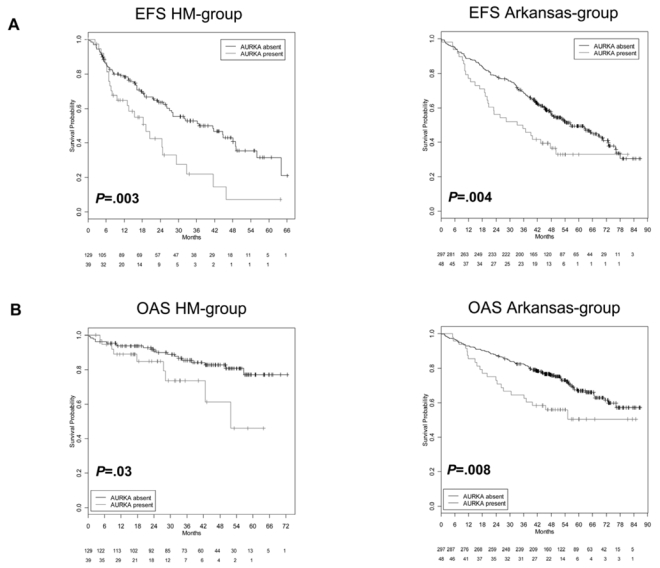
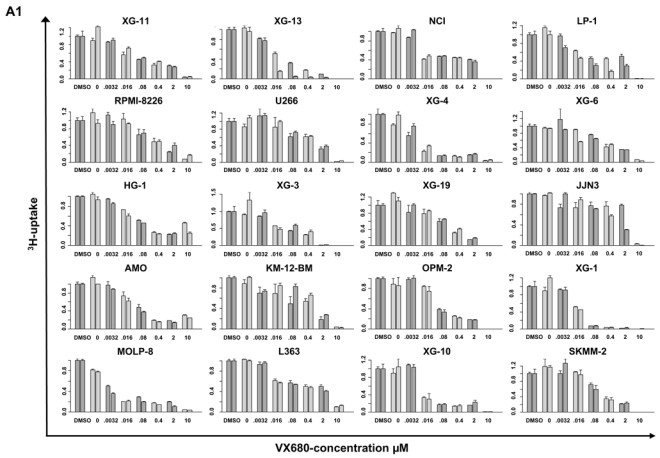
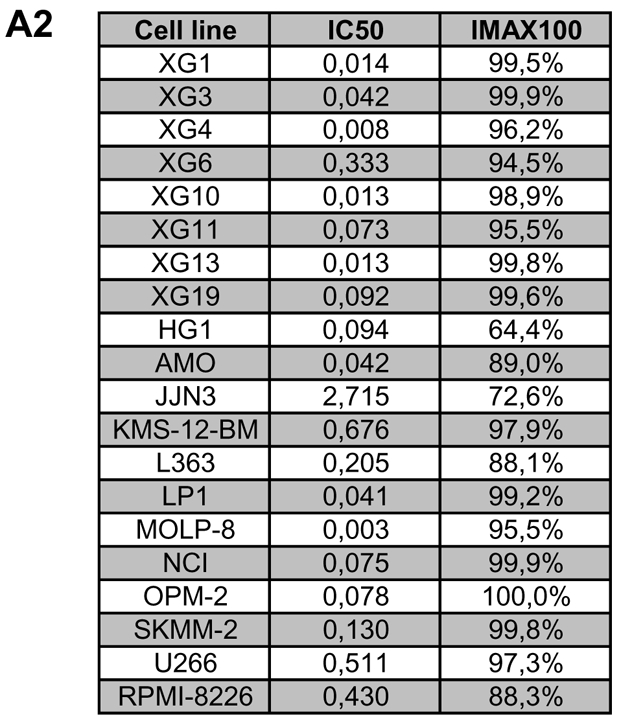
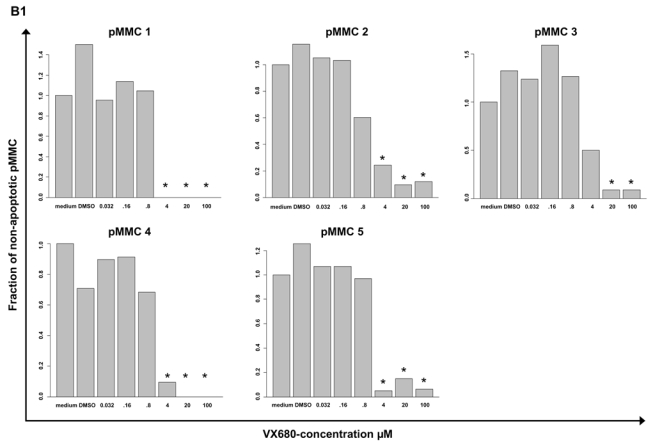
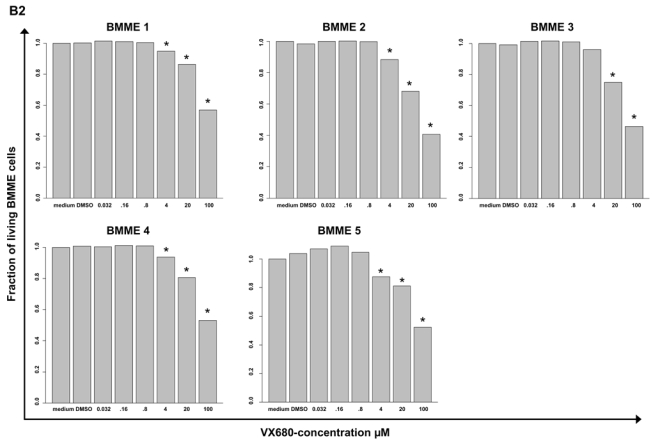
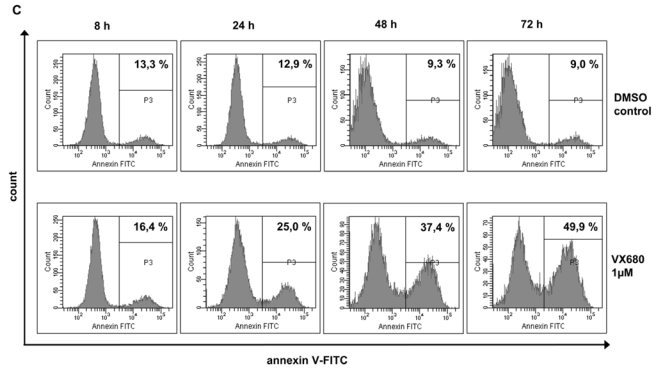
Genetic instability and cellular proliferation have been associated with aurora kinase expression in several cancer entities, including multiple myeloma. Therefore, the expression of aurora-A, -B, and -C was determined by Affymetrix DNA microarrays in 784 samples including 2 independent sets of 233 and 345 CD138-purified myeloma cells from previously untreated patients. Chromosomal aberrations were assessed by comprehensive interphase fluorescence in situ hybridization and proliferation of primary myeloma cells by propidium iodine staining. We found aurora-A and -B to be expressed at varying frequencies in primary myeloma cells of different patient cohorts, but aurora-C in testis cell samples only. Myeloma cell samples with detectable versus absent aurora-A expression show a significantly higher proliferation rate, but neither a higher absolute number of chromosomal aberrations (aneuploidy), nor of subclonal aberrations (chromosomal instability). The clinical aurora kinase inhibitor VX680 induced apoptosis in 20 of 20 myeloma cell lines and 5 of 5 primary myeloma cell samples. Presence of aurora-A expression delineates significantly inferior event-free and overall survival in 2 independent cohorts of patients undergoing high-dose chemotherapy, independent from conventional prognostic factors. Using gene expression profiling, aurora kinase inhibitors as a promising therapeutic option in myeloma can be tailoredly given to patients expressing aurora-A, who in turn have an adverse prognosis.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Similar articles
-
Proliferation is a central independent prognostic factor and target for personalized and risk-adapted treatment in multiple myeloma.Haematologica. 2011 Jan;96(1):87-95. doi: 10.3324/haematol.2010.030296. Epub 2010 Sep 30. Haematologica. 2011. PMID: 20884712 Free PMC article.
-
Aurora A kinase RNAi and small molecule inhibition of Aurora kinases with VE-465 induce apoptotic death in multiple myeloma cells.Leuk Lymphoma. 2008 Mar;49(3):559-69. doi: 10.1080/10428190701824544. Leuk Lymphoma. 2008. PMID: 18297535
-
The selective Aurora B kinase inhibitor AZD1152 is a potential new treatment for multiple myeloma.Br J Haematol. 2008 Feb;140(3):295-302. doi: 10.1111/j.1365-2141.2007.06913.x. Epub 2007 Dec 10. Br J Haematol. 2008. PMID: 18076711
-
Aurora kinase inhibitors as anticancer molecules.Biochim Biophys Acta. 2010 Oct-Dec;1799(10-12):829-39. doi: 10.1016/j.bbagrm.2010.09.004. Epub 2010 Sep 20. Biochim Biophys Acta. 2010. PMID: 20863917 Free PMC article. Review.
-
Aurora B: a new prognostic marker and therapeutic target in cancer.Curr Med Chem. 2011;18(4):482-96. doi: 10.2174/092986711794480203. Curr Med Chem. 2011. PMID: 21143115 Review.
Cited by
-
Genome instability in multiple myeloma.Leukemia. 2020 Nov;34(11):2887-2897. doi: 10.1038/s41375-020-0921-y. Epub 2020 Jul 10. Leukemia. 2020. PMID: 32651540 Review.
-
Bortezomib reduces the tumorigenicity of multiple myeloma via downregulation of upregulated targets in clonogenic side population cells.PLoS One. 2013;8(3):e56954. doi: 10.1371/journal.pone.0056954. Epub 2013 Mar 4. PLoS One. 2013. PMID: 23469177 Free PMC article.
-
Matrine and CYC116 synergistically inhibit growth and induce apoptosis in multiple myeloma cells.Chin J Integr Med. 2015 Aug;21(8):635-9. doi: 10.1007/s11655-015-1975-y. Epub 2015 Mar 24. Chin J Integr Med. 2015. PMID: 25804197
-
Preclinical efficacy of sepantronium bromide (YM155) in multiple myeloma is conferred by down regulation of Mcl-1.Oncotarget. 2014 Nov 15;5(21):10237-50. doi: 10.18632/oncotarget.2529. Oncotarget. 2014. PMID: 25296978 Free PMC article.
-
An interactive nomogram based on clinical and molecular signatures to predict prognosis in multiple myeloma patients.Aging (Albany NY). 2021 Jul 14;13(14):18442-18463. doi: 10.18632/aging.203294. Epub 2021 Jul 14. Aging (Albany NY). 2021. PMID: 34260414 Free PMC article.
References
-
- Kyle RA, Rajkumar SV. Multiple myeloma. N Engl J Med. 2004;351:1860–1873. - PubMed
-
- Witzig TE, Timm M, Larson D, Therneau T, Greipp PR. Measurement of apoptosis and proliferation of bone marrow plasma cells in patients with plasma cell proliferative disorders. Br J Haematol. 1999;104:131–137. - PubMed
-
- Boccadoro M, Gavarotti P, Fossati G, et al. Low plasma cell 3(H) thymidine incorporation in monoclonal gammopathy of undetermined significance (MGUS), smouldering myeloma and remission phase myeloma: a reliable indicator of patients not requiring therapy. Br J Haematol. 1984;58:689–696. - PubMed
-
- Greipp PR, Lust JA, O’Fallon WM, et al. Plasma cell labeling index and beta 2-microglobulin predict survival independent of thymidine kinase and C-reactive protein in multiple myeloma. Blood. 1993;81:3382–3387. - PubMed
-
- Cremer FW, Bila J, Buck I, et al. Delineation of distinct subgroups of multiple myeloma and a model for clonal evolution based on interphase cytogenetics. Genes Chromosomes Cancer. 2005;44:194–203. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
