

doi: 10.1038/nchembio.147.
Epub 2009 Jan 26.
Discovering chemical modifiers of oncogene-regulated hematopoietic differentiation
Affiliations
- PMID: 19172146
- PMCID: PMC2658727
- DOI: 10.1038/nchembio.147
Item in Clipboard
Discovering chemical modifiers of oncogene-regulated hematopoietic differentiation
Nat Chem Biol.
2009 Apr.
Abstract
It has been proposed that inhibitors of an oncogene's effects on multipotent hematopoietic progenitor cell differentiation may change the properties of the leukemic stem cells and complement the clinical use of cytotoxic drugs. Using zebrafish, we developed a robust in vivo hematopoietic differentiation assay that reflects the activity of the oncogene AML1-ETO. Screening for modifiers of AML1-ETO-mediated hematopoietic dysregulation uncovered unexpected roles of COX-2- and beta-catenin-dependent pathways in AML1-ETO function. This approach may open doors for developing therapeutics targeting oncogene function within leukemic stem cells.
Figures
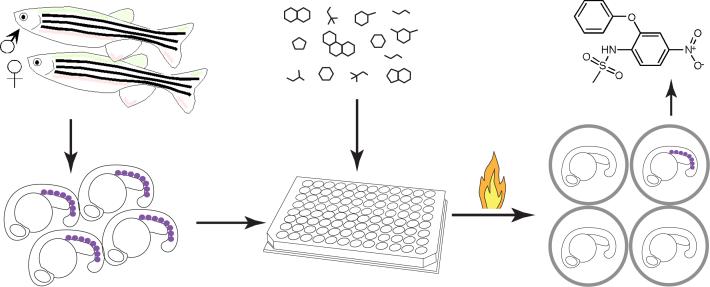
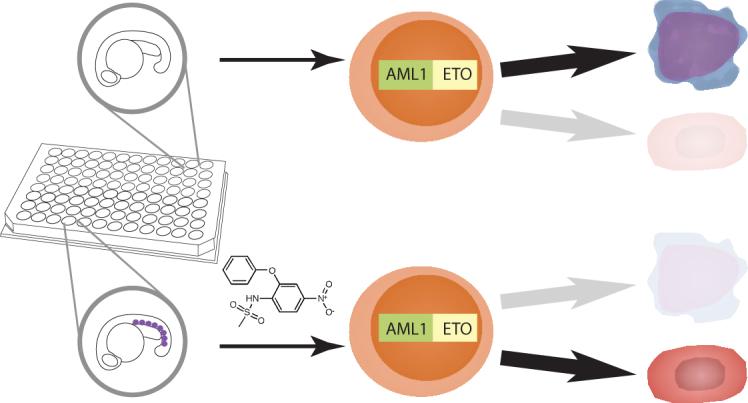
Screening for chemical suppressors of AE. Homozygous Tg(hsp:AML1-ETO) fish were crossed with wild-type fish to generate thousands of heterozygous Tg(hsp:AML1-ETO) embryos. These embryos were raised for 12−16 hours post fertilization at which point five embryos were arrayed into each well of the 96-well plates. The compounds from the library were added to the plates. An hour later, the plates were heat-shocked at 40 °C for 1 hour to induce AE expression. At 90 minutes after the heat shock, the embryos were processed for in situ hybridization of gata1. Induced expression of AE resulted in lost of gata1+ hematopoietic cells (indicated as purple dots) in the posterior blood islands of the zebrafish embryos. However, the chemical suppressors of AE, such as nimesulide (shown in the figure), antagonized AE's effect, restoring gata1 expression in Tg(hsp:AML1-ETO) embryos.
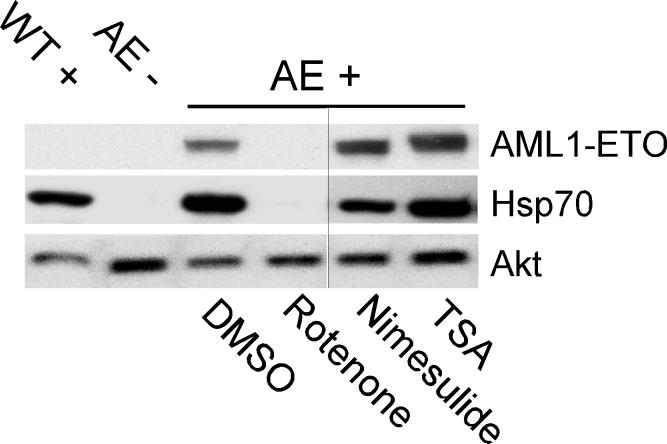
Nimesulide does not affect AE expression in Tg(hsp:AML1-ETO) embryos. Western blot analysis shows that while rotenone (0.15 μM) inhibits heat-induced expression of AE and Hsp70, neither nimesulide (40 μM) nor trichostatin A (TSA, 0.5 μM) affects heat-induced expression of AE and Hsp70. The expression of Akt is used as a reference for protein loading. At the concentrations used, all three compounds suppress the zebrafish AE phenotype. DMSO is used as the vehicle control. WT+, wild-type embryos with heat treatment. AE-, Tg(hsp:AML1-ETO) embryos without heat treatment. AE+, Tg(hsp:AML1-ETO) embryos with heat treatment. The vertical black line indicates the juxtaposition of lanes that were not contiguous in the original gel.
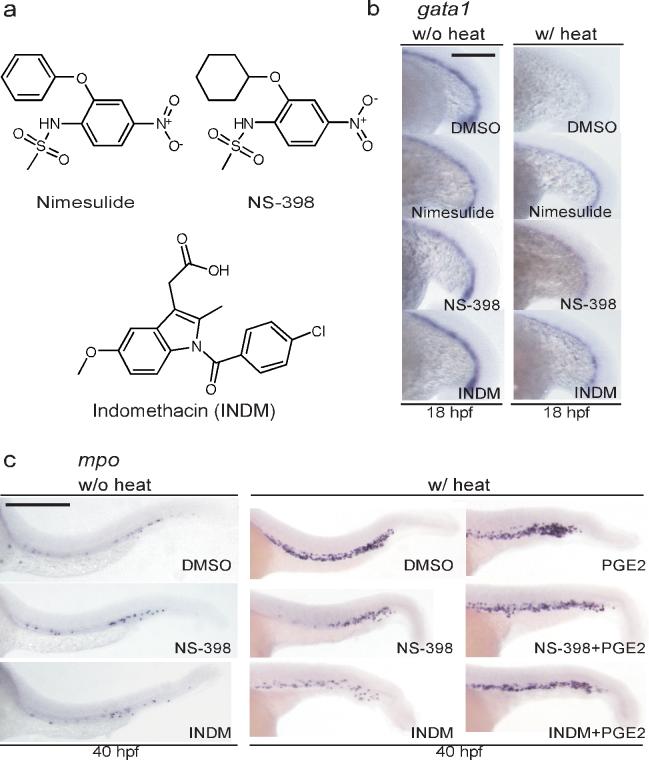
Cyclooxygenase (COX) inhibitors reverse AE's effects on hematopoietic differentiation. (a) The chemical structures of the selective COX-2 inhibitors nimesulide, NS-398, and the non-selective COX inhibitor indomethacin (INDM). (b) In situ hybridization of gata1. The COX inhibitors restore gata1 expression in Tg(hsp:AML1-ETO) embryos. To induce AE expression, Tg(hsp:AML1-ETO) embryos were incubated at 39 °C for 1 hour at the 16-somite stage (w/ heat). Induced expression of AE results in the loss of gata1 expression as compared to Tg(hsp:AML1-ETO) embryos without heat incubation. Nevertheless, addition of nimesulide (40 μM), NS-398 (25 μM) or INDM (20 μM) reverses AE's effects. All compounds were added to the embryos 1 hour before the heat shock. DMSO was used as the vehicle control. Scale bar, 0.3 mm. (c) In situ hybridization of mpo. NS-398 (25 μM) and INDM (20 μM) antagonize AE's effect, suppressing the accumulation of mpo+ granulocytic cells. The antagonism no longer exists in the presence of prostaglandin E2 (20 μM). All compounds were added at the end of heat shock. Scale bar, 0.3 mm.
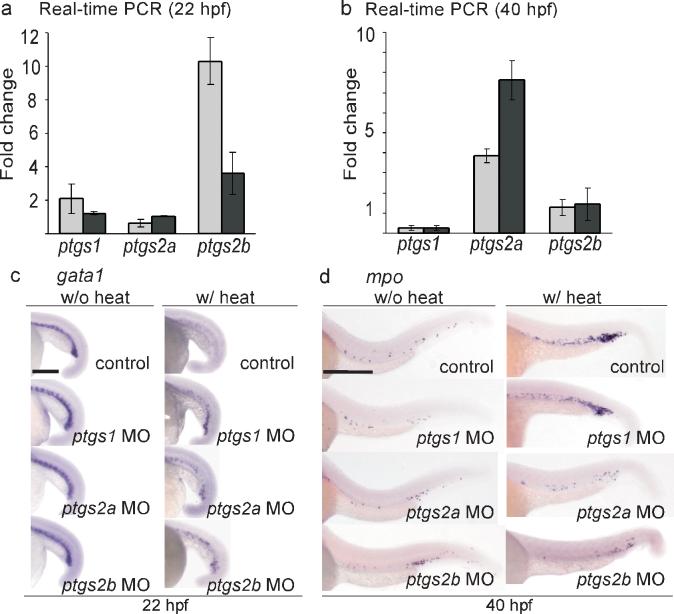
The hematopoietic phenotype of AE involves induction of the genes that encode both zebrafish COX-2 isoforms. (a) Real-time PCR indicates that ptgs2b is upregulated in the hematopoietic cells of Tg(hsp:AML1-ETO) embryos at 2 hours after heat shock. Total RNA was extracted from the hematopoietic cells isolated from heat-treated wild-type and Tg(hsp:AML1-ETO) embryos at 22 hpf. The expression of ptgs1, ptgs2a, and ptgs2b was evaluated by real-time PCR analysis using two independent primer sets for each gene, and was normalized to gapdh expression. The fold change indicates the ratio of mRNA expression between Tg(hsp:AML1-ETO) and wild-type embryos. ptgs1, 2.09±0.0.91, 1.22±0.08; ptgs2a, 0.62±0.21; 1.04±0.03; ptgs2b, 10.31±1.39, 3.60±1.25 (mean±SEM). (b) Real-time PCR indicates that ptgs2a is upregulated in the hematopoietic cells of Tg(hsp:AML1-ETO) embryos at 40 hpf. ptgs1, 0.26±0.12, 0.28±0.11; ptgs2a, 3.86±0.36; 7.6±0.98; ptgs2b, 1.28±0.39, 1.45±0.81 (mean±SEM). (c) In situ hybridization of gata1. Knockdown of ptgs1, ptgs2a, or ptgs2b partially restored gata1 expression in heat-treated Tg(hsp:AML1-ETO) embryos. Scale bar, 0.3 mm. (d) In situ hybridization of mpo. Knockdown of ptgs2a or ptgs2b suppresses AE-induced mpo expression. Control, non-injected embryos. Scale bar, 0.3 mm.
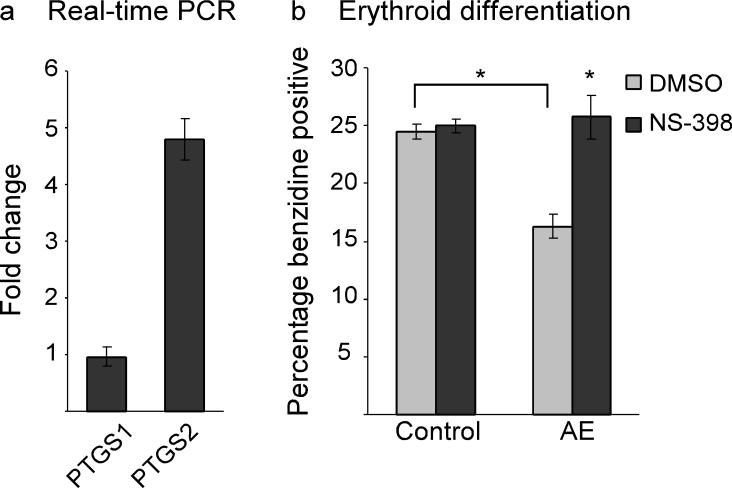
Erythroid differentiation of human myelogenous leukemia K562 cells is attenuated by AE via a COX-2-dependent mechanism. (a) Real-time PCR indicates that expression of AE causes upregulation of PTGS2 but not PTGS1 in K562 cells. K562 cells containing stable integration of the vector pLRT-GFP (clone B9) or the AE expression vector pLRT-AE (clone D8) were harvested for RNA extraction. The expression of PTGS1 and PTGS2 was evaluated by real-time PCR analysis and normalized to 18S RNA expression. The fold change indicates the ratio of mRNA expression between D8 and B9 cells. PTGS1, 0.962±0.169; PTGS2, 4.798±0.373 (mean±SEM). (b) Erythroid differentiation efficiency as scored by benzidine staining. While AE suppresses erythroid differentiation of K562 cells, inhibition of COX-2 by NS-398 (75 μM) reverses AE's effect. K562 clone B9 (Control) and clone D8 (AE) were treated with either DMSO or NS-398 for 4 days before staining. Control-DMSO, 24.5±0.65; Control-NS-398, 25±0.58; AE-DMSO, 16.25±1.03; AE-NS-398, 25.75±1.89 (mean±SEM). *p<0.01 (two-tailed t-test).
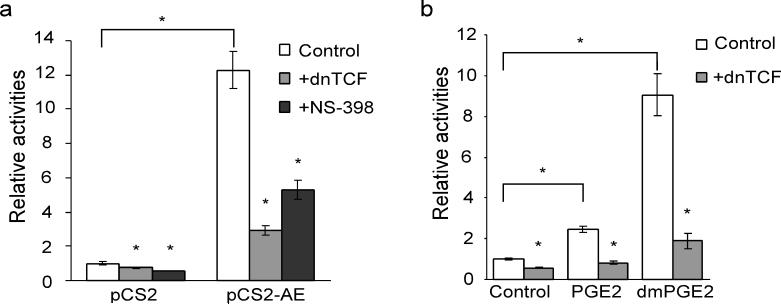
AE activates β-catenin-dependent transcription through COX-2. (a) Expression of AE induces TOPflash luciferase activity, which can be suppressed by overexpression of a dominant-negative form of TCF (dnTCF) or by NS-398 (75 μM). The empty vector (pCS2) or the expression vector for AE (pCS2-AE) was transfected into K562 cells along with TOPflash and pRL-tk which encodes Renilla luciferase for normalizing the transfection efficiency. The results are shown in relative TOPflash luciferase activities after normalization. pCS2/control, 1±0.09; pCS2+dnTCF, 0.73±0.03; pCS2+NS-398, 0.58±0.01; pCS2-AE/control, 12.29±1.09, pCS2-AE+dnTCF,2.93±0.3; pCS2-AE+NS-398, 5.29±0.57 (mean±SEM). *p<0.01 (two-tailed t-test). (b) 16,16-dimethylprostaglandin E2 (dmPGE2) activates β-catenin-TCF-dependent transcription. K562 cells were transfected with TOPflash and pRL-tk. At 4 hours after transfection, PGE2 (20 μM), dmPGE2 (20 μg ml−1) or DMSO was added to the cells. The luciferase activities were measured two days after the transfection. In addition, the dmPGE2-induced TOPflash activity can be suppressed by overexpression of dnTCF. control, 1±0.05; control+dn-TCF, 0.56±0.03; PGE2, 2.44±0.15; PGE2+dnTCF, 0.83±0.1; dmPGE2, 9.06±1.02; dmPGE2+dnTCF, 1.89±0.38 (mean±SEM). *p<0.01 (two-tailed t-test).
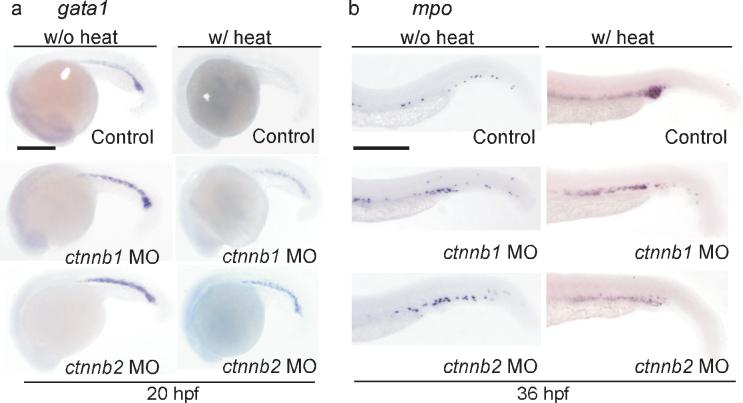
The hematopoietic differentiation effects caused by AE are dependent on β-catenin function. (a-b) In situ hybridization of gata1 (a) and mpo (b). Knockdown of either β-catenin1 ctnnb1 MO) or β-catenin2 (ctnnb2 MO) restores gata1 expression and suppresses mpo upregulation in the presence of AE. Control, non-injected embryos. Scale bar, 0.3 mm.
Similar articles
-
AML1-ETO reprograms hematopoietic cell fate by downregulating scl expression.Development. 2008 Jan;135(2):401-10. doi: 10.1242/dev.008904. Development. 2008. PMID: 18156164
-
AML1-ETO mediates hematopoietic self-renewal and leukemogenesis through a COX/β-catenin signaling pathway.Blood. 2013 Jun 13;121(24):4906-16. doi: 10.1182/blood-2012-08-447763. Epub 2013 May 3. Blood. 2013. PMID: 23645839 Free PMC article.
-
Zebrafish small molecule screen in reprogramming/cell fate modulation.Methods Mol Biol. 2010;636:317-27. doi: 10.1007/978-1-60761-691-7_20. Methods Mol Biol. 2010. PMID: 20336532 Free PMC article.
-
Oncogenic pathways of AML1-ETO in acute myeloid leukemia: multifaceted manipulation of marrow maturation.Cancer Lett. 2007 Jun 28;251(2):179-86. doi: 10.1016/j.canlet.2006.10.010. Epub 2006 Nov 27. Cancer Lett. 2007. PMID: 17125917 Free PMC article. Review.
-
Effects of the leukemia-associated AML1-ETO protein on hematopoietic stem and progenitor cells.Oncogene. 2004 May 24;23(24):4249-54. doi: 10.1038/sj.onc.1207673. Oncogene. 2004. PMID: 15156180 Review.
Cited by
-
From phenotype to mechanism after zebrafish small molecule screens.Drug Discov Today Dis Models. 2013 Spring;10(1):e51-e55. doi: 10.1016/j.ddmod.2012.02.002. Drug Discov Today Dis Models. 2013. PMID: 26146505 Free PMC article.
-
Identification of mundoserone by zebrafish in vivo screening as a natural product with anti-angiogenic activity.Exp Ther Med. 2018 Dec;16(6):4562-4568. doi: 10.3892/etm.2018.6748. Epub 2018 Sep 17. Exp Ther Med. 2018. PMID: 30542405 Free PMC article.
-
Zebrafish and Medaka: new model organisms for modern biomedical research.J Biomed Sci. 2016 Jan 28;23:19. doi: 10.1186/s12929-016-0236-5. J Biomed Sci. 2016. PMID: 26822757 Free PMC article. Review.
-
Development of an In Vitro Assay to Quantitate Hematopoietic Stem and Progenitor Cells (HSPCs) in Developing Zebrafish Embryos.J Vis Exp. 2017 Nov 30;(129):56836. doi: 10.3791/56836. J Vis Exp. 2017. PMID: 29286381 Free PMC article.
-
RUNX1 and RUNX1-ETO: roles in hematopoiesis and leukemogenesis.Front Biosci (Landmark Ed). 2012 Jan 1;17(3):1120-39. doi: 10.2741/3977. Front Biosci (Landmark Ed). 2012. PMID: 22201794 Free PMC article. Review.
References
-
- Redaelli A, Botteman MF, Stephens JM, Brandt S, Pashos CL. Economic burden of acute myeloid leukemia: a literature review. Cancer Treat Rev. 2004;30:237–247. - PubMed
-
- Guan Y, Gerhard B, Hogge DE. Detection, isolation, and stimulation of quiescent primitive leukemic progenitor cells from patients with acute myeloid leukemia (AML). Blood. 2003;101:3142–3149. - PubMed
-
- Terpstra W, et al. Fluorouracil selectively spares acute myeloid leukemia cells with long-term growth abilities in immunodeficient mice and in culture. Blood. 1996;88:1944–1950. - PubMed
-
- Wang JC, Dick JE. Cancer stem cells: lessons from leukemia. Trends Cell Biol. 2005;15:494–501. - PubMed
-
- Tenen DG. Disruption of differentiation in human cancer: AML shows the way. Nat Rev Cancer. 2003;3:89–101. - PubMed
Publication types
MeSH terms
Substances
Associated data
- PubChem-Substance/56459045
- PubChem-Substance/56459046
- PubChem-Substance/56459047
- PubChem-Substance/56459048
- PubChem-Substance/56459049
- PubChem-Substance/56459050
- PubChem-Substance/56459051
- PubChem-Substance/56459052
- PubChem-Substance/56459053
- PubChem-Substance/56459054
- PubChem-Substance/56459055
- PubChem-Substance/56459056
- PubChem-Substance/56459057
- PubChem-Substance/56459058
- PubChem-Substance/56459059
- PubChem-Substance/56459060
- PubChem-Substance/56459061
- PubChem-Substance/56459062
- PubChem-Substance/56459063
- PubChem-Substance/56459064
- PubChem-Substance/56459065
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
