

Chromatin condensation in terminally differentiating mouse erythroblasts does not involve special architectural proteins but depends on histone deacetylation
- PMID: 19172406
- PMCID: PMC2667965
- DOI: 10.1007/s10577-008-9005-y
Chromatin condensation in terminally differentiating mouse erythroblasts does not involve special architectural proteins but depends on histone deacetylation
Abstract
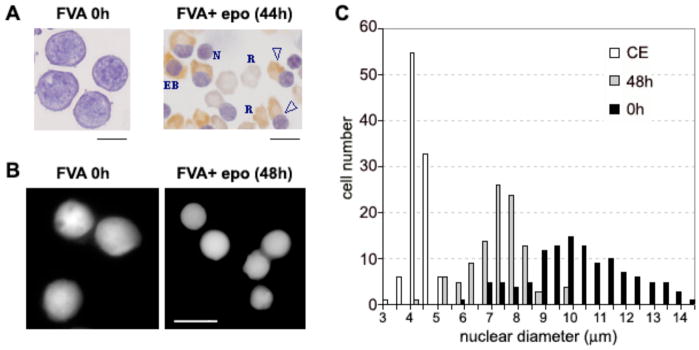
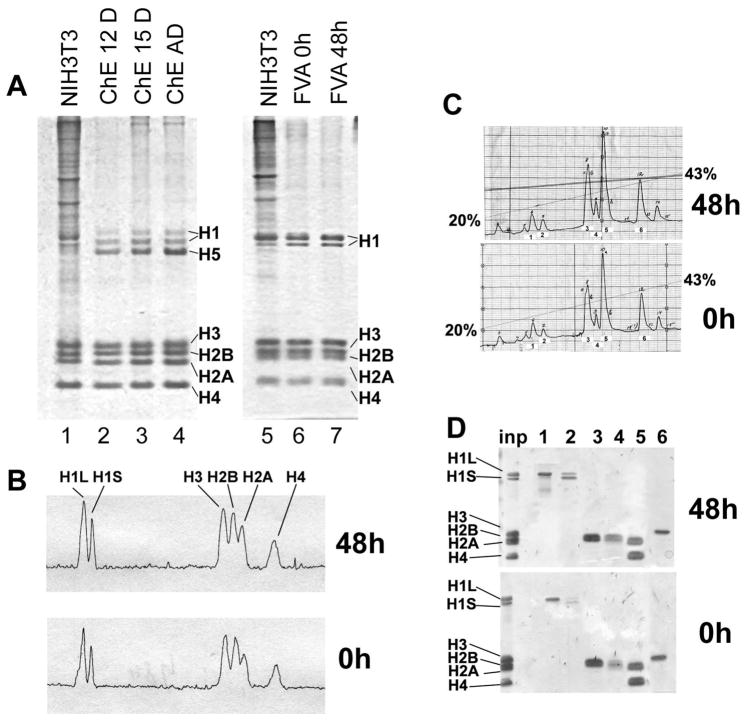
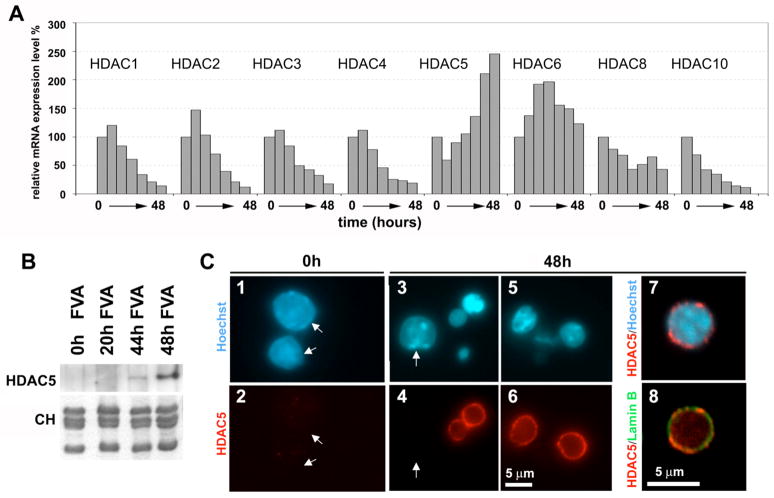
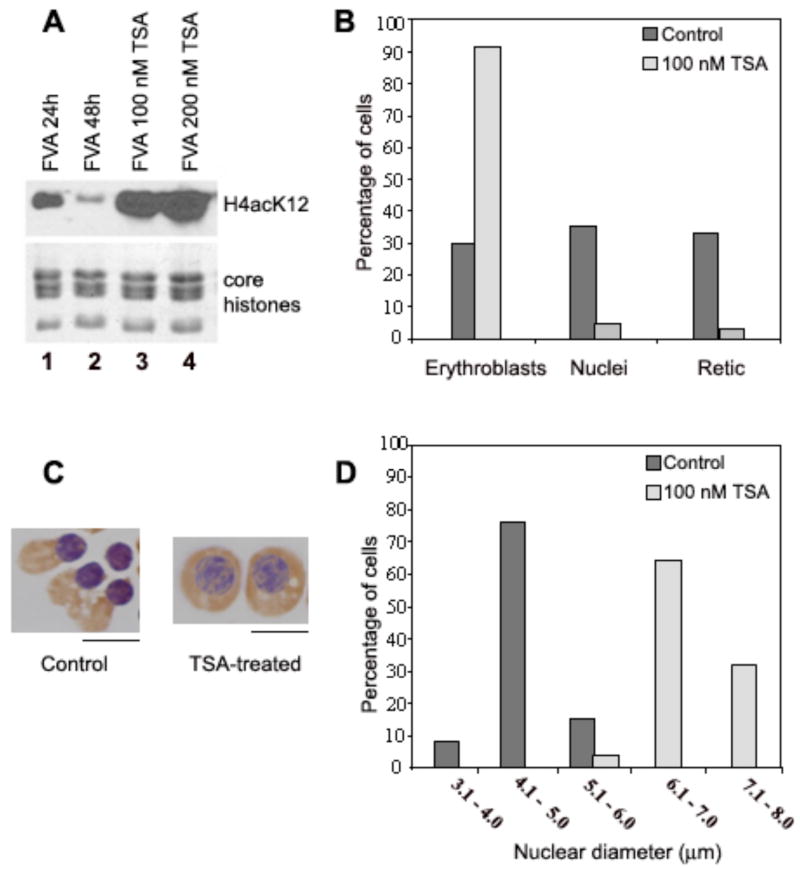
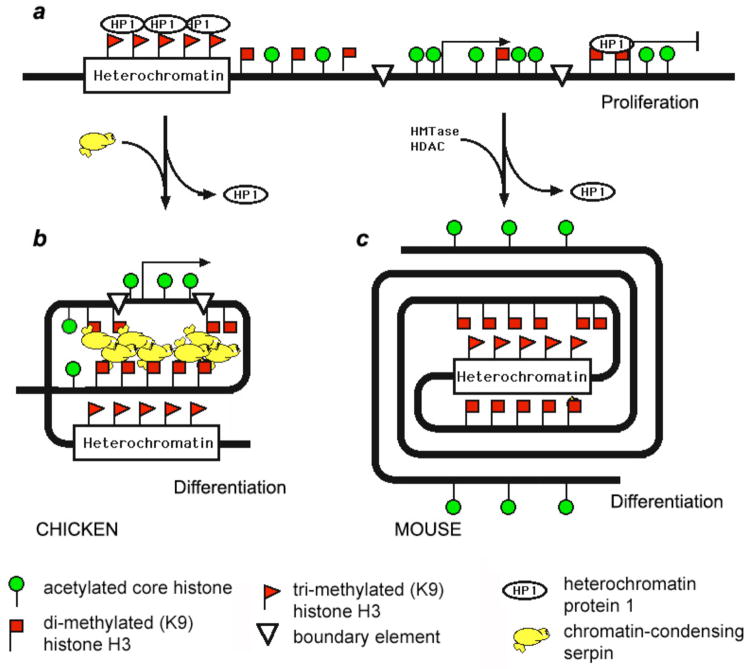
Terminal erythroid differentiation in vertebrates is characterized by progressive heterochromatin formation and chromatin condensation and, in mammals, culminates in nuclear extrusion. To date, although mechanisms regulating avian erythroid chromatin condensation have been identified, little is known regarding this process during mammalian erythropoiesis. To elucidate the molecular basis for mammalian erythroblast chromatin condensation, we used Friend virus-infected murine spleen erythroblasts that undergo terminal differentiation in vitro. Chromatin isolated from early and late-stage erythroblasts had similar levels of linker and core histones, only a slight difference in nucleosome repeats, and no significant accumulation of known developmentally regulated architectural chromatin proteins. However, histone H3(K9) dimethylation markedly increased while histone H4(K12) acetylation dramatically decreased and became segregated from the histone methylation as chromatin condensed. One histone deacetylase, HDAC5, was significantly upregulated during the terminal stages of Friend virus-infected erythroblast differentiation. Treatment with histone deacetylase inhibitor, trichostatin A, blocked both chromatin condensation and nuclear extrusion. Based on our data, we propose a model for a unique mechanism in which extensive histone deacetylation at pericentromeric heterochromatin mediates heterochromatin condensation in vertebrate erythroblasts that would otherwise be mediated by developmentally-regulated architectural proteins in nucleated blood cells.
Figures



References
-
- Adkins NL, Watts M, Georgel PT. To the 30-nm chromatin fiber and beyond. Biochim Biophys Acta. 2004;1677:12–23. - PubMed
-
- Bannister AJ, Schneider R, Myers FA, Thorne AW, Crane-Robinson C, Kouzarides T. Spatial distribution of Di- and tri-methyl lysine 36 of histone H3 at active genes. J Biol Chem. 2005;280:17732–17736. - PubMed
-
- Bernstein E, Allis CD. RNA meets chromatin. Genes Dev. 2005;19:1635–1655. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
