

Essential role of c-Cbl in amphiregulin-induced recycling and signaling of the endogenous epidermal growth factor receptor
- PMID: 19173594
- PMCID: PMC2645952
- DOI: 10.1021/bi801771g
Essential role of c-Cbl in amphiregulin-induced recycling and signaling of the endogenous epidermal growth factor receptor
Abstract
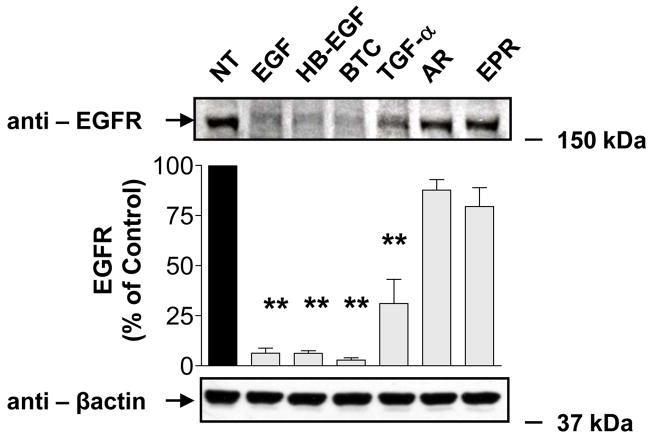
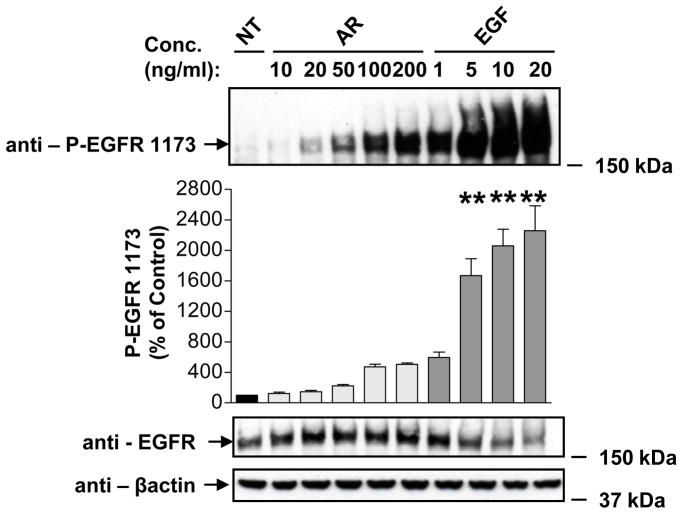
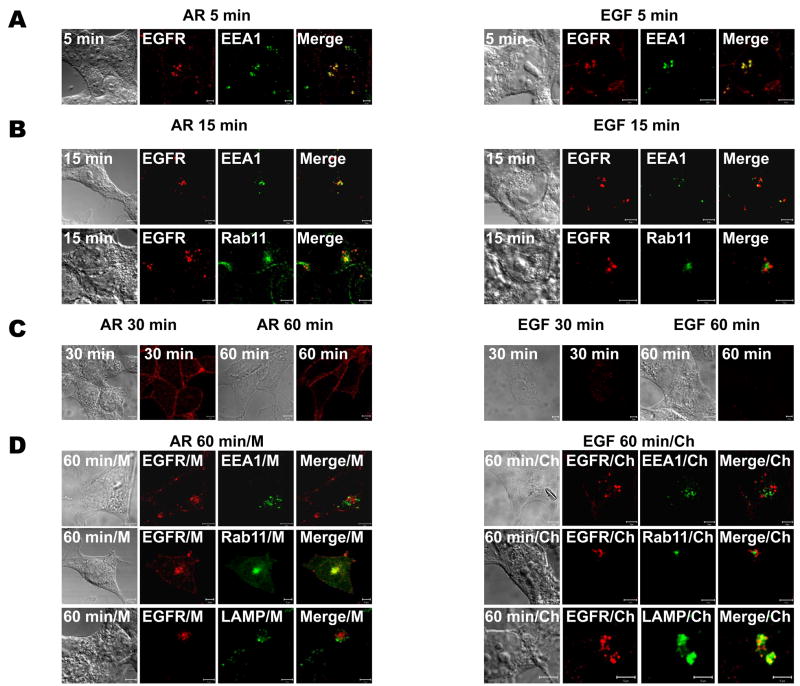
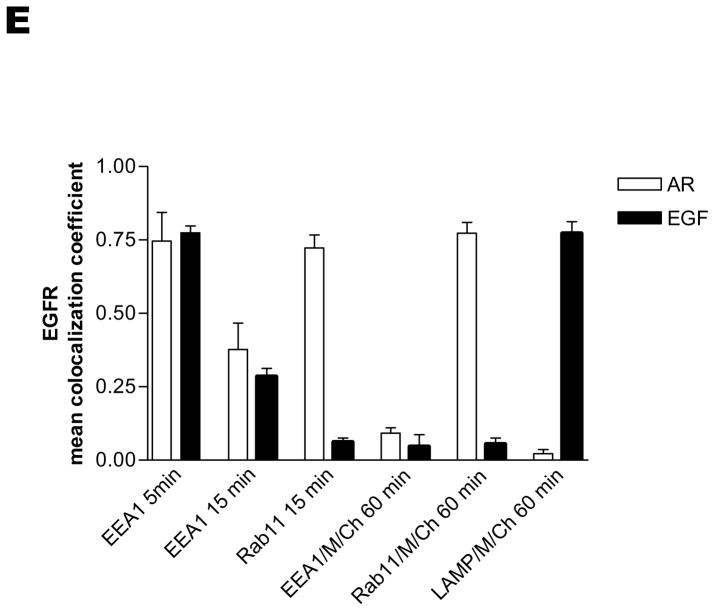
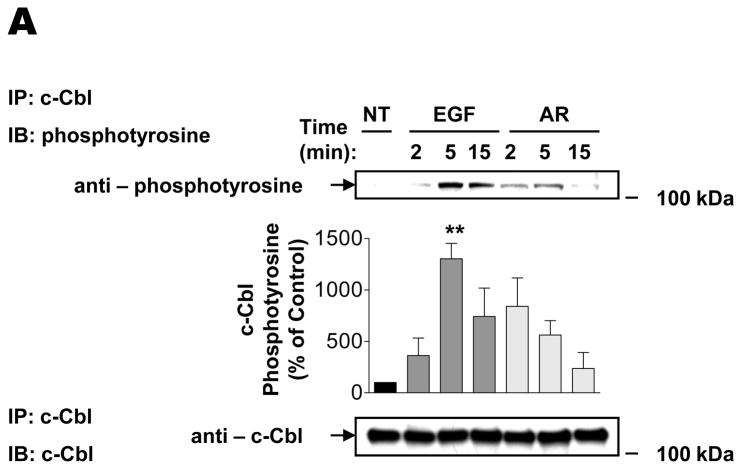
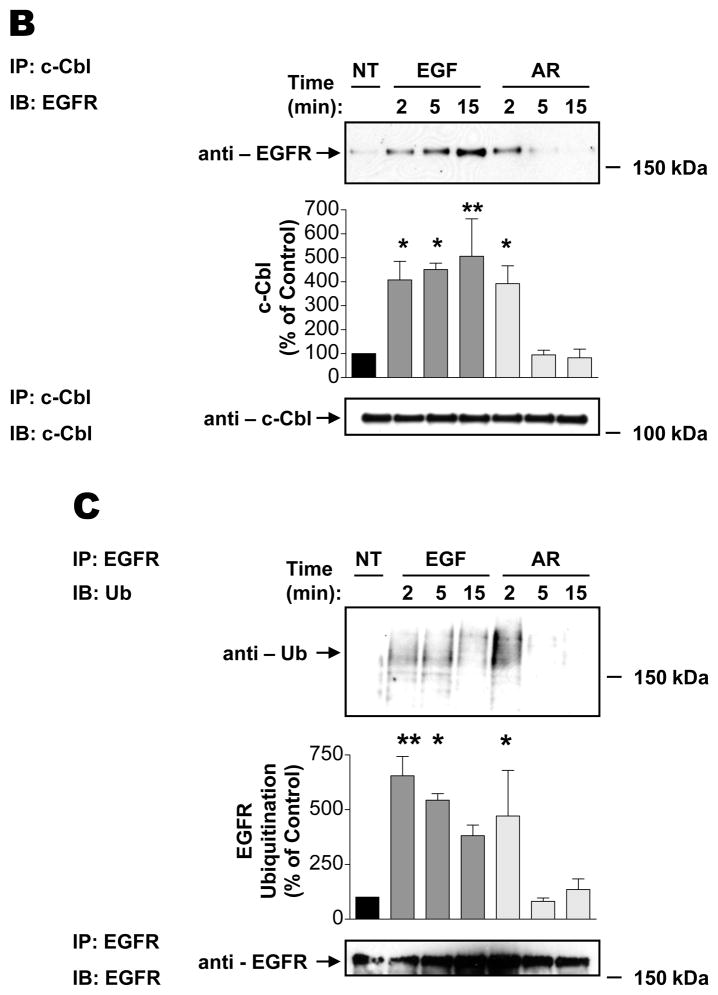
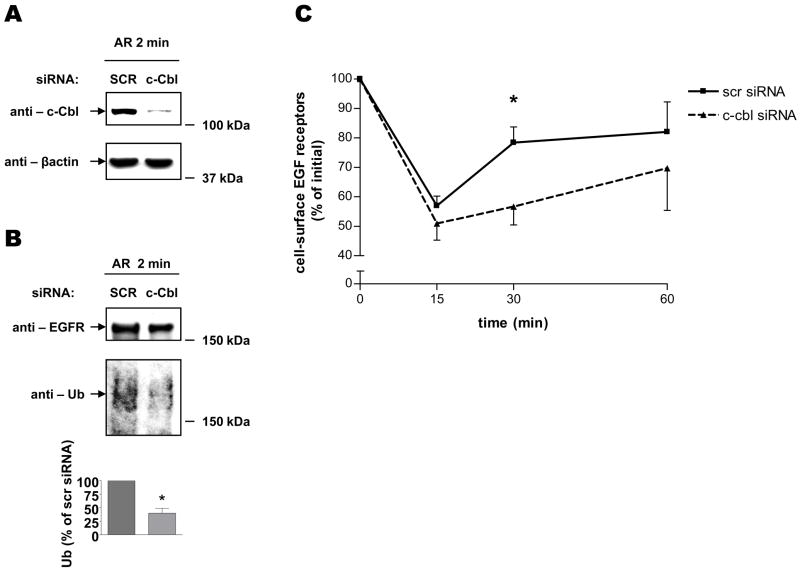
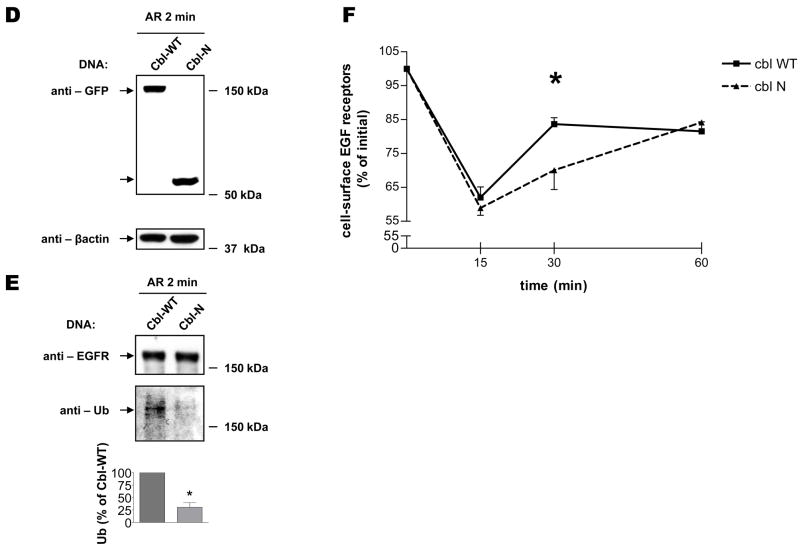
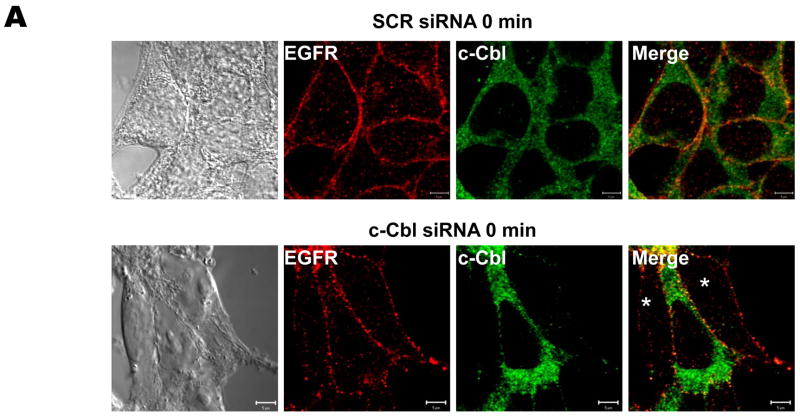
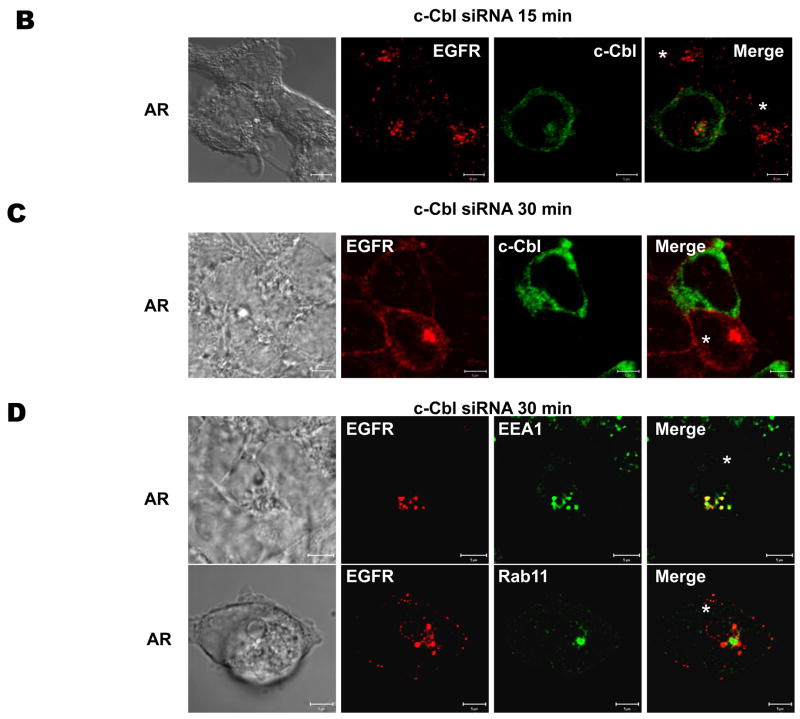
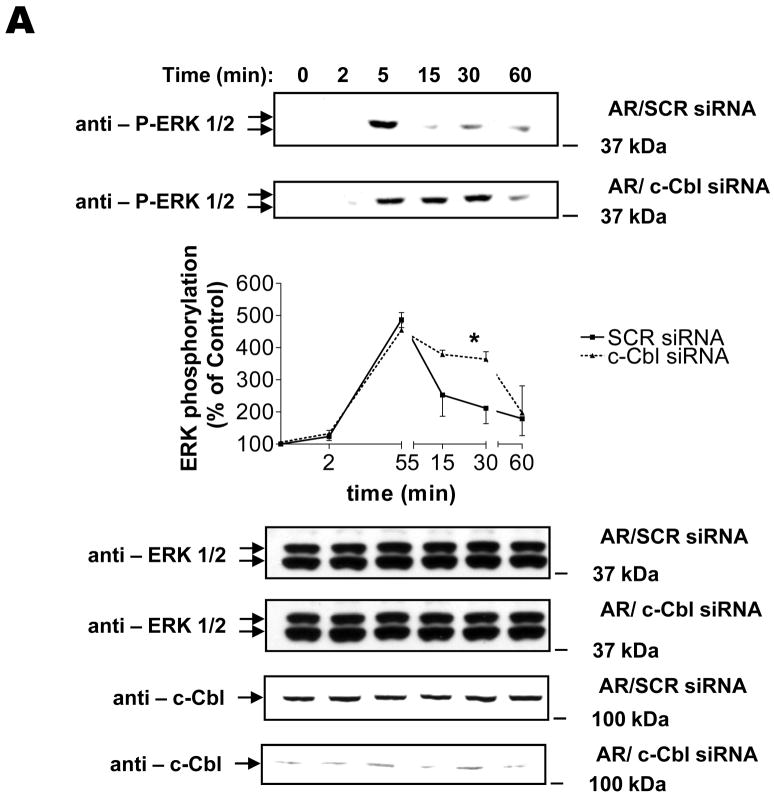
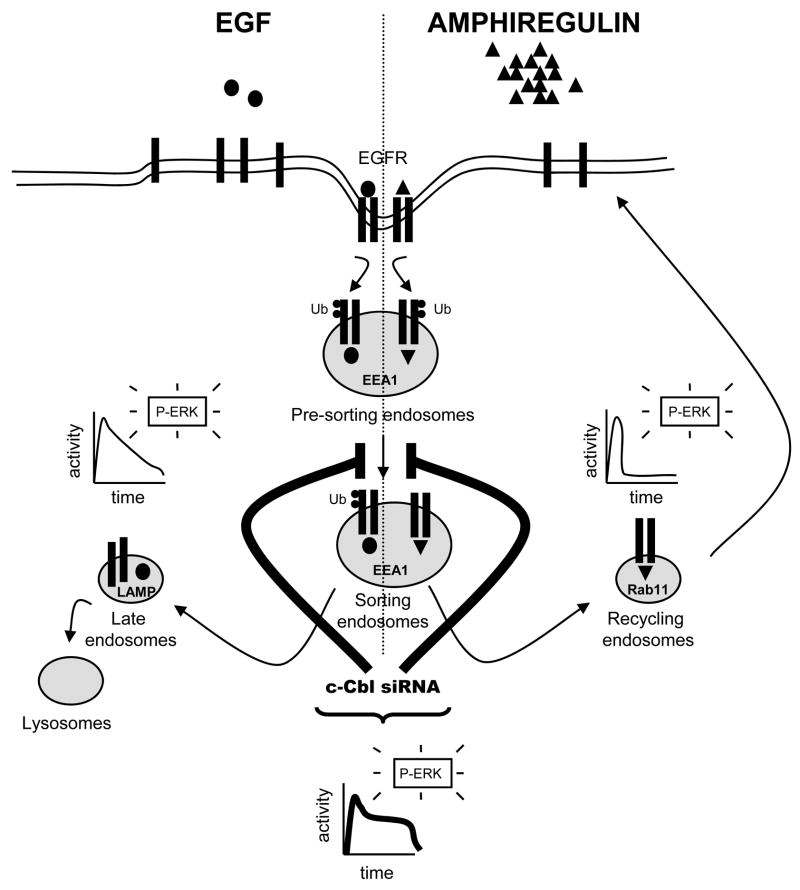
The intracellular processing of the epidermal growth factor receptor (EGFR) induced by epidermal growth factor (EGF) and transforming growth factor-alpha (TGF-alpha) has been studied meticulously, with the former resulting in EGFR degradation and the latter in EGFR recycling to the plasma membrane. However, little is known about how other EGF family growth factors affect the trafficking of the EGFR. Additionally, although both EGF and TGF-alpha have been shown to effectively induce initial c-Cbl (ubiquitin ligase)-mediated ubiquitination of the EGFR, limited information is available regarding the role of c-Cblin the trafficking and signaling of recycling EGFR. Thus, in this study, we investigated the roles of c-Cblin endogenous EGFR trafficking and signaling after stimulation with amphiregulin (AR). We demonstrated that a physiological concentration of AR induced recycling of the endogenous EGFR to the plasma membrane, which correlated closely with transient association of the EGFR with c-Cbl and transient EGFR ubiquitination. Most importantly, we used c-Cbl small interfering RNA (siRNA) duplexes and ac-Cbl dominant negative mutant to show that c-Cbl is critical for the efficient transition of the EGFR from early endosomes to a recycling pathway and that c-Cbl regulates the duration of extracellular signal regulated kinase 1/2 mitogen-activated protein kinase (ERK1/2 MAPK) phosphorylation. These data support novel functions of c-Cbl in mediating recycling of EGF receptors to the plasma membrane, as well as in mediating the duration of activation (transient vs sustained) of ERK1/2 MAPK phosphorylation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
