

Presenilin 1 affects focal adhesion site formation and cell force generation via c-Src transcriptional and posttranslational regulation
- PMID: 19176482
- PMCID: PMC2665068
- DOI: 10.1074/jbc.M806825200
Presenilin 1 affects focal adhesion site formation and cell force generation via c-Src transcriptional and posttranslational regulation
Abstract
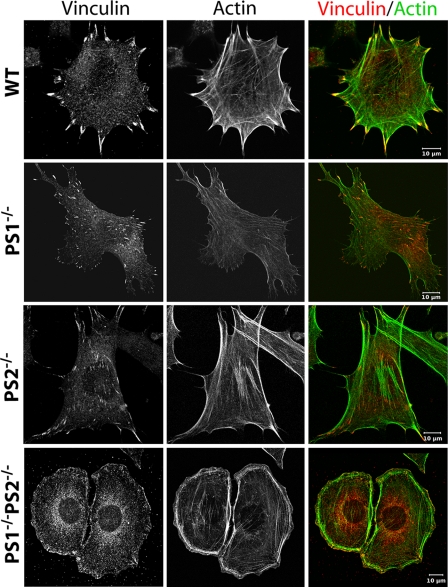
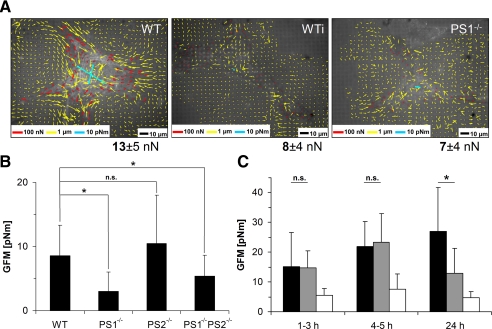
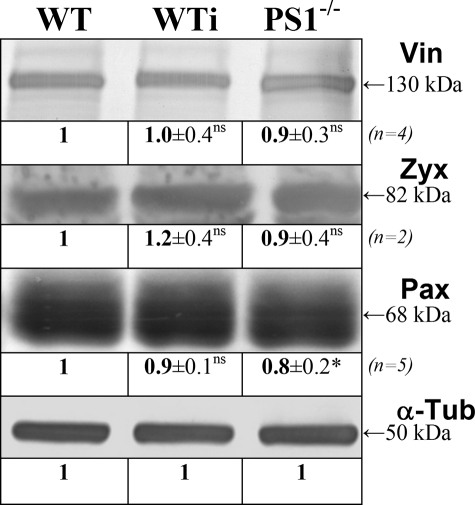
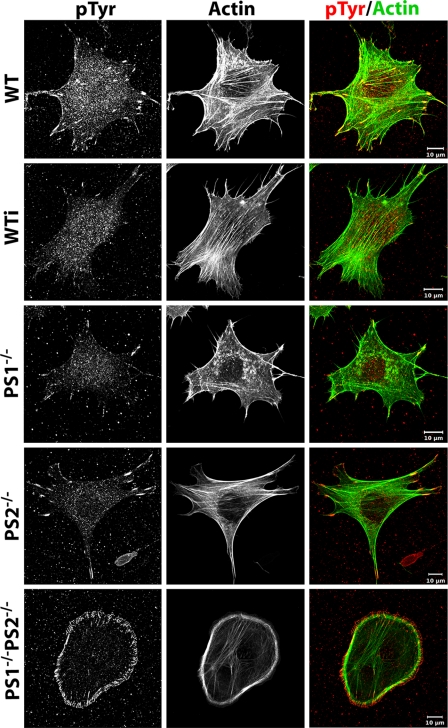
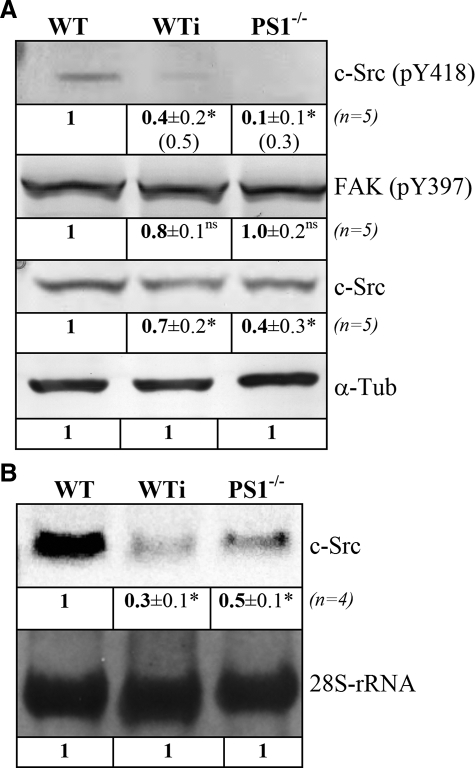
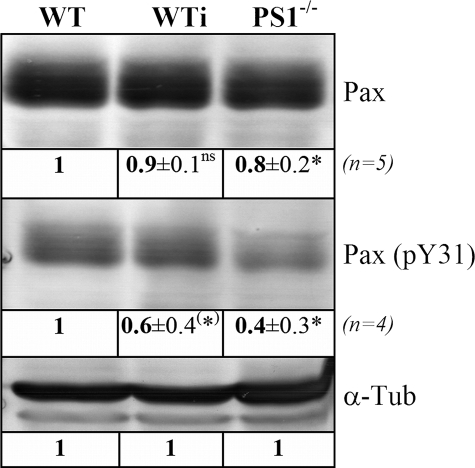
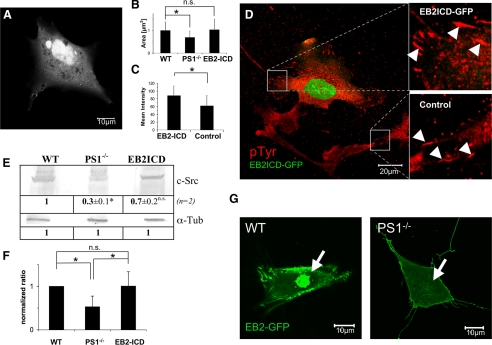
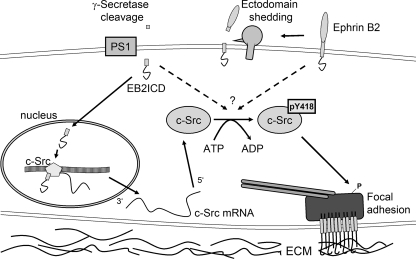
Presenilin 1 and 2 (PS) are critical components of the gamma-secretase complex that cleaves type I transmembrane proteins within their transmembrane domains. This process leads to release of proteolytically processed products from cellular membranes and plays an essential role in signal transduction or vital functions as cell adhesion. Here we studied the function of presenilins in cell-matrix interaction of wild-type and PS knock-out mouse embryonic fibroblasts. We found for PS1(-/-) cells an altered morphology with significantly reduced sizes of focal adhesion sites compared with wild type. Cell force analyses on micropatterned elastomer films revealed PS1(-/-) cell forces to be reduced by 50%. Pharmacological inhibition confirmed this function of gamma-secretase in adhesion site and cell force formation. On the regulatory level, PS1 deficiency was associated with strongly decreased phosphotyrosine levels of focal adhesion site-specific proteins. The reduced tyrosine phosphorylation was caused by a down-regulation of c-Src kinase activity primarily at the level of c-Src transcription. The direct regulatory connection between PS1 and c-Src could be identified with ephrinB2 as PS1 target protein. Overexpression of ephrinB2 cytoplasmic domain resulted in its nuclear translocation with increased levels of c-Src and a full complementation of the PS1(-/-) adhesion and phosphorylation phenotype. Cleavage of full-length EB2 and subsequent intracellular domain translocation depended on PS1 as these processes were only found in WT cells. Therefore, we conclude that gamma-secretase is vital for controlling cell adhesion and force formation by transcriptional regulation of c-Src via ephrinB2 cleavage.
Figures

References
-
- De Strooper, B., Saftig, P., Craessaerts, K., Vanderstichele, H., Guhde, G., Annaert, W., Von Figura, K., and Van Leuven, F. (1998) Nature 391 387-390 - PubMed
-
- De Strooper, B., Annaert, W., Cupers, P., Saftig, P., Craessaerts, K., Mumm, J. S., Schroeter, E. H., Schrijvers, V., Wolfe, M. S., Ray, W. J., Goate, A., and Kopan, R. (1999) Nature 398 518-522 - PubMed
-
- Xia, W., and Wolfe, M. S. (2003) J. Cell Sci. 116 2839-2844 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
