

Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis
- PMID: 19176721
- PMCID: PMC2649400
- DOI: 10.1104/pp.108.130369
Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis
Abstract
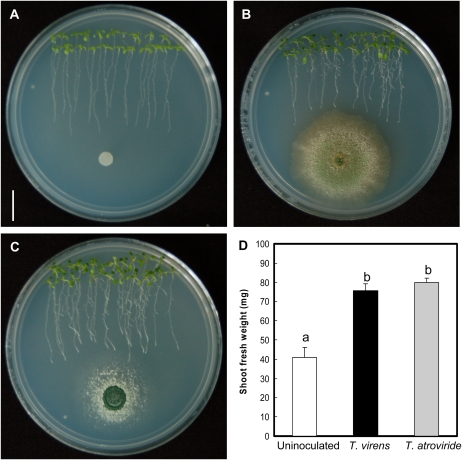
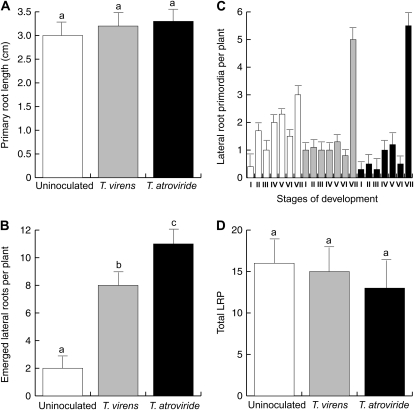
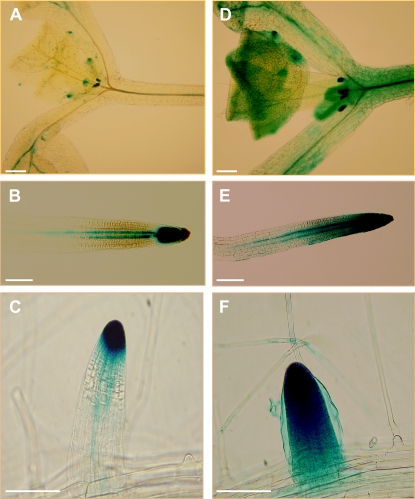
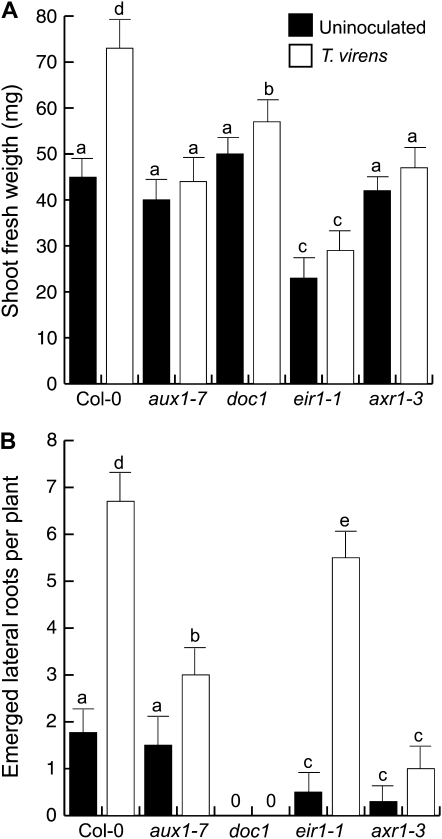
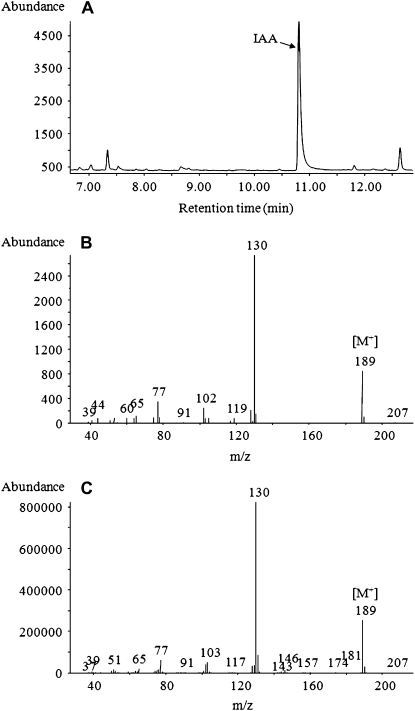
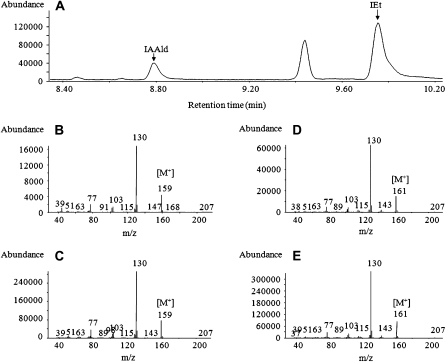
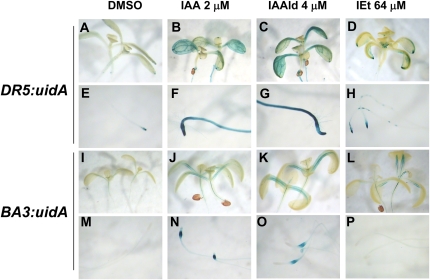
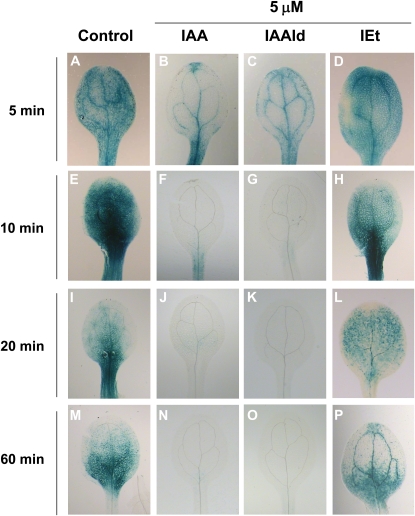
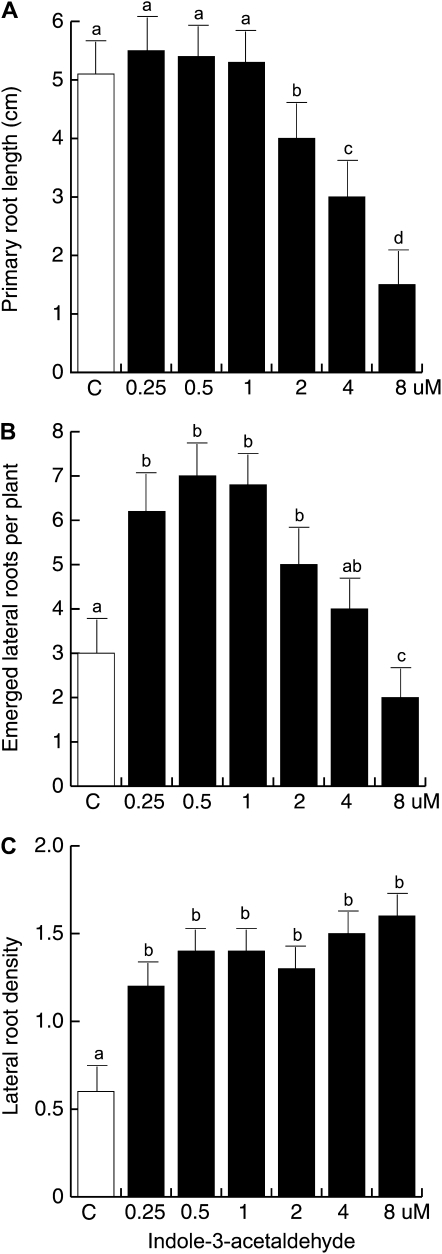
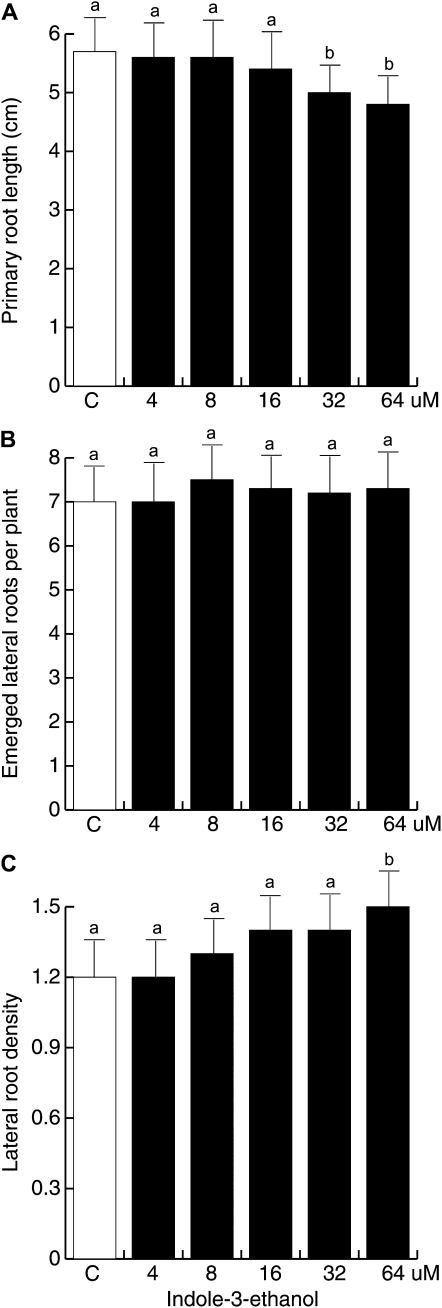
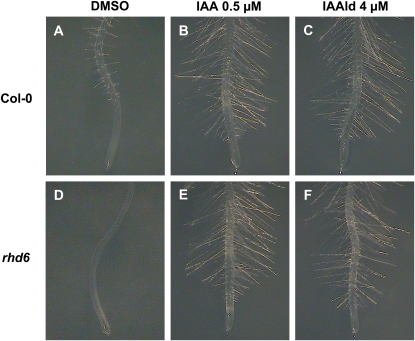
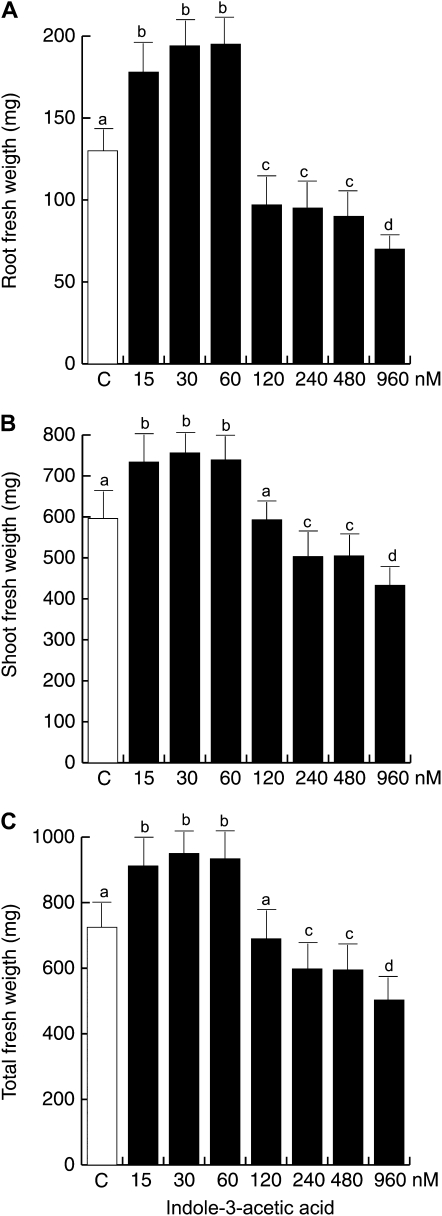
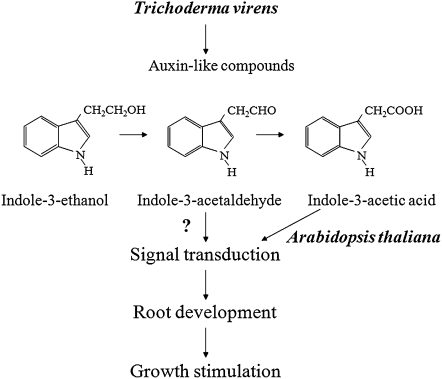
Trichoderma species belong to a class of free-living fungi beneficial to plants that are common in the rhizosphere. We investigated the role of auxin in regulating the growth and development of Arabidopsis (Arabidopsis thaliana) seedlings in response to inoculation with Trichoderma virens and Trichoderma atroviride by developing a plant-fungus interaction system. Wild-type Arabidopsis seedlings inoculated with either T. virens or T. atroviride showed characteristic auxin-related phenotypes, including increased biomass production and stimulated lateral root development. Mutations in genes involved in auxin transport or signaling, AUX1, BIG, EIR1, and AXR1, were found to reduce the growth-promoting and root developmental effects of T. virens inoculation. When grown under axenic conditions, T. virens produced the auxin-related compounds indole-3-acetic acid, indole-3-acetaldehyde, and indole-3-ethanol. A comparative analysis of all three indolic compounds provided detailed information about the structure-activity relationship based on their efficacy at modulating root system architecture, activation of auxin-regulated gene expression, and rescue of the root hair-defective phenotype of the rhd6 auxin response Arabidopsis mutant. Our results highlight the important role of auxin signaling for plant growth promotion by T. virens.
Figures
Similar articles
-
Trichoderma spp. Improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na⁺ elimination through root exudates.Mol Plant Microbe Interact. 2014 Jun;27(6):503-14. doi: 10.1094/MPMI-09-13-0265-R. Mol Plant Microbe Interact. 2014. PMID: 24502519
-
Mitogen-Activated Protein Kinase 6 and Ethylene and Auxin Signaling Pathways Are Involved in Arabidopsis Root-System Architecture Alterations by Trichoderma atroviride.Mol Plant Microbe Interact. 2015 Jun;28(6):701-10. doi: 10.1094/MPMI-01-15-0005-R. Epub 2015 Jun 12. Mol Plant Microbe Interact. 2015. PMID: 26067203
-
The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning.New Phytol. 2016 Mar;209(4):1496-512. doi: 10.1111/nph.13725. Epub 2015 Nov 16. New Phytol. 2016. PMID: 26568541
-
Mechanisms for plant growth promotion activated by Trichoderma in natural and managed terrestrial ecosystems.Microbiol Res. 2024 Apr;281:127621. doi: 10.1016/j.micres.2024.127621. Epub 2024 Jan 18. Microbiol Res. 2024. PMID: 38295679 Review.
-
Auxin and the integration of environmental signals into plant root development.Ann Bot. 2013 Dec;112(9):1655-65. doi: 10.1093/aob/mct229. Epub 2013 Oct 17. Ann Bot. 2013. PMID: 24136877 Free PMC article. Review.
Cited by
-
Dual functionality of Trichoderma: Biocontrol of Sclerotinia sclerotiorum and biostimulant of cotton plants.Front Plant Sci. 2022 Oct 7;13:983127. doi: 10.3389/fpls.2022.983127. eCollection 2022. Front Plant Sci. 2022. PMID: 36275544 Free PMC article.
-
Photosynthetic electron transport rate and root dynamics of finger millet in response to Trichoderma harzianum.Plant Signal Behav. 2022 Dec 31;17(1):2146373. doi: 10.1080/15592324.2022.2146373. Plant Signal Behav. 2022. PMID: 36382615 Free PMC article.
-
Trichoderma: The "Secrets" of a Multitalented Biocontrol Agent.Plants (Basel). 2020 Jun 18;9(6):762. doi: 10.3390/plants9060762. Plants (Basel). 2020. PMID: 32570799 Free PMC article. Review.
-
Bacterial endophytes as indicators of susceptibility to Cercospora Leaf Spot (CLS) disease in Beta vulgaris L.Sci Rep. 2022 Jun 23;12(1):10719. doi: 10.1038/s41598-022-14769-8. Sci Rep. 2022. PMID: 35739218 Free PMC article.
-
Efficacy of biological agents and fillers seed coating in improving drought stress in anise.Front Plant Sci. 2022 Jul 22;13:955512. doi: 10.3389/fpls.2022.955512. eCollection 2022. Front Plant Sci. 2022. PMID: 35937352 Free PMC article.
References
-
- Adams P, De-Leij FA, Lynch JM (2007) Trichoderma harzianum Rifai 1295-22 mediates growth promotion of Crack willow (Salix fragilis) saplings in both clean and metal-contaminated soil. Microb Ecol 54 306–313 - PubMed
-
- Bais HP, Weir TL, Perry L, Gilroy S, Vivanco JM (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annu Rev Plant Biol 57 233–266 - PubMed
-
- Bjorkman T, Blanchard LM, Harman GE (1998) Growth enhancement of shrunken-2 sweet corn when colonized with Trichoderma harzianum 1295-22: effect of environmental stress. J Am Soc Hortic Sci 123 35–40
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
