

Backbone structure of a small helical integral membrane protein: A unique structural characterization
- PMID: 19177358
- PMCID: PMC2708045
- DOI: 10.1002/pro.24
Backbone structure of a small helical integral membrane protein: A unique structural characterization
Abstract
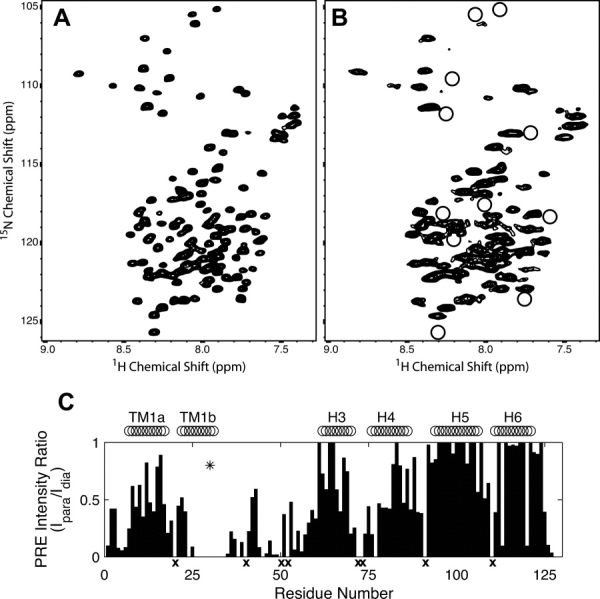
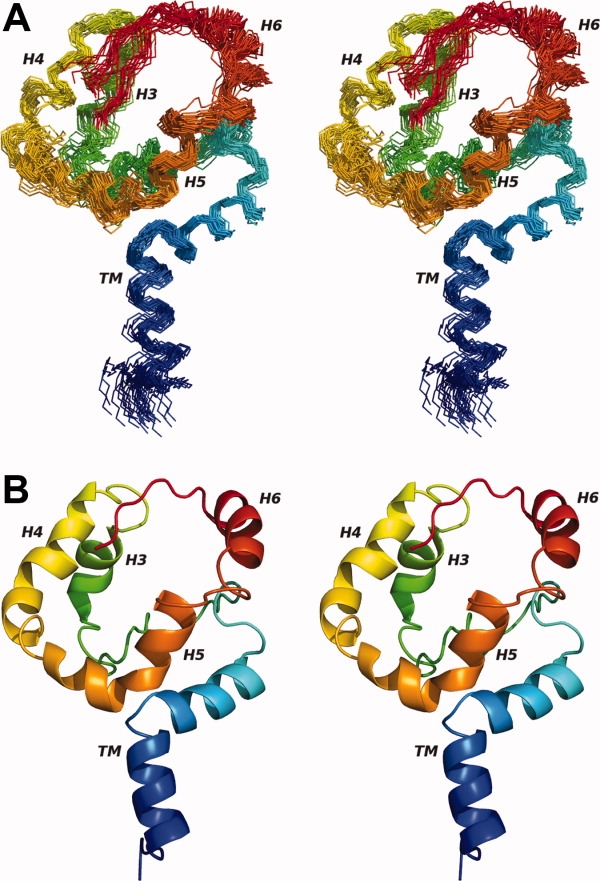
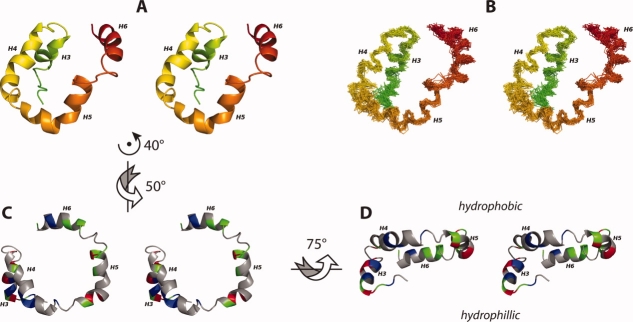
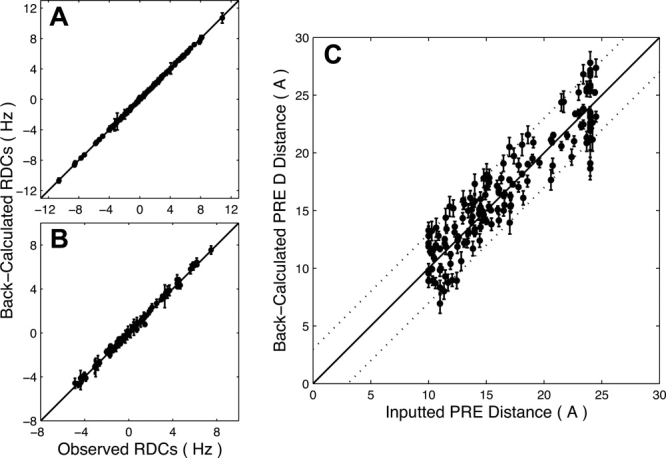
The structural characterization of small integral membrane proteins pose a significant challenge for structural biology because of the multitude of molecular interactions between the protein and its heterogeneous environment. Here, the three-dimensional backbone structure of Rv1761c from Mycobacterium tuberculosis has been characterized using solution NMR spectroscopy and dodecylphosphocholine (DPC) micelles as a membrane mimetic environment. This 127 residue single transmembrane helix protein has a significant (10 kDa) C-terminal extramembranous domain. Five hundred and ninety distance, backbone dihedral, and orientational restraints were employed resulting in a 1.16 A rmsd backbone structure with a transmembrane domain defined at 0.40 A. The structure determination approach utilized residual dipolar coupling orientation data from partially aligned samples, long-range paramagnetic relaxation enhancement derived distances, and dihedral restraints from chemical shift indices to determine the global fold. This structural model of Rv1761c displays some influences by the membrane mimetic illustrating that the structure of these membrane proteins is dictated by a combination of the amino acid sequence and the protein's environment. These results demonstrate both the efficacy of the structural approach and the necessity to consider the biophysical properties of membrane mimetics when interpreting structural data of integral membrane proteins and, in particular, small integral membrane proteins.
Figures

References
-
- Nishimura K, Kim S, Zhang L, Cross TA. The closed state of a H+ channel helical bundle combining precise orientational and distance restraints from solid state NMR. Biochemistry. 2002;41:13170–13177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
