

Chemokines in the immunopathogenesis of hepatitis C infection
- PMID: 19177577
- PMCID: PMC2919201
- DOI: 10.1002/hep.22763
Chemokines in the immunopathogenesis of hepatitis C infection
Abstract
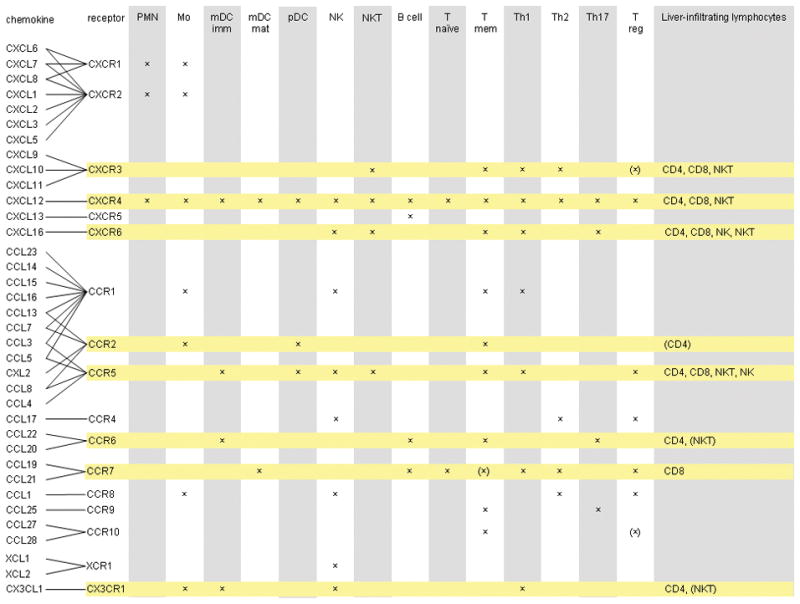
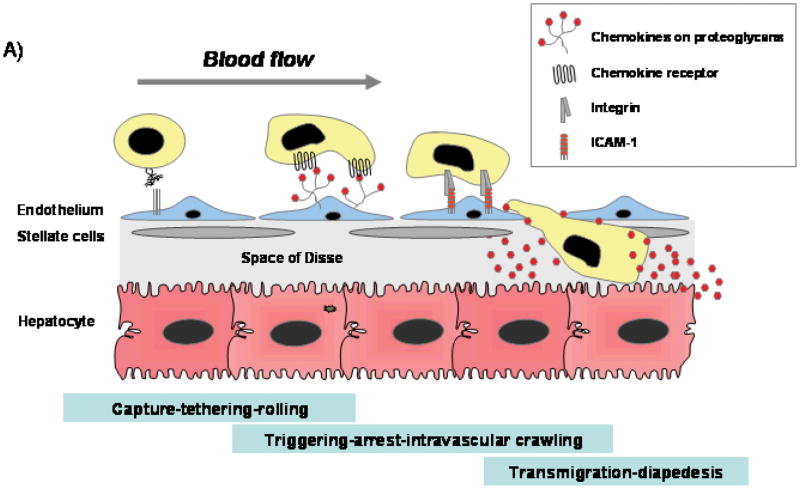
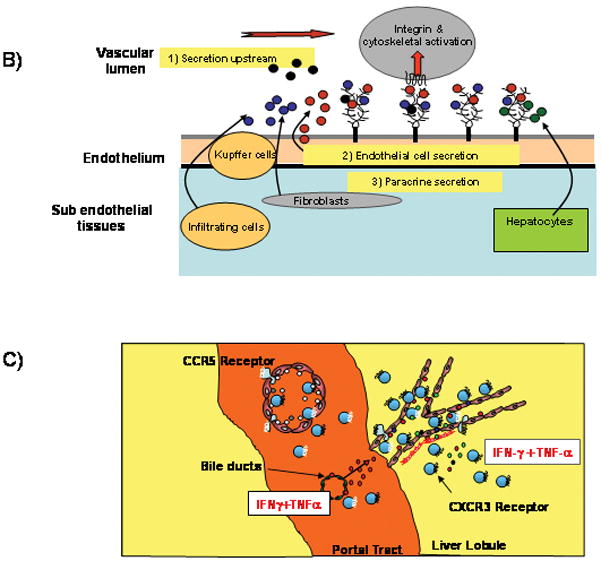
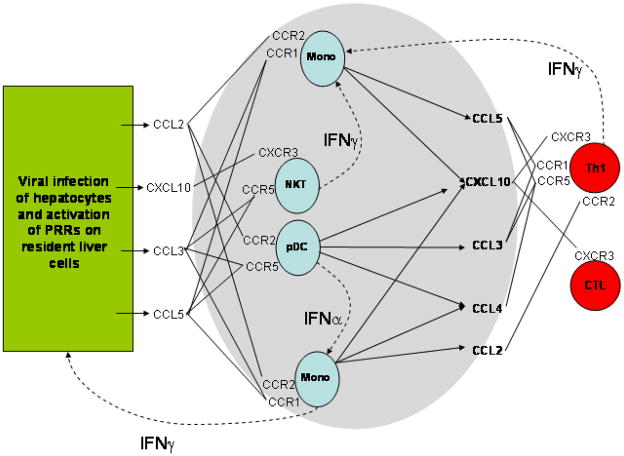
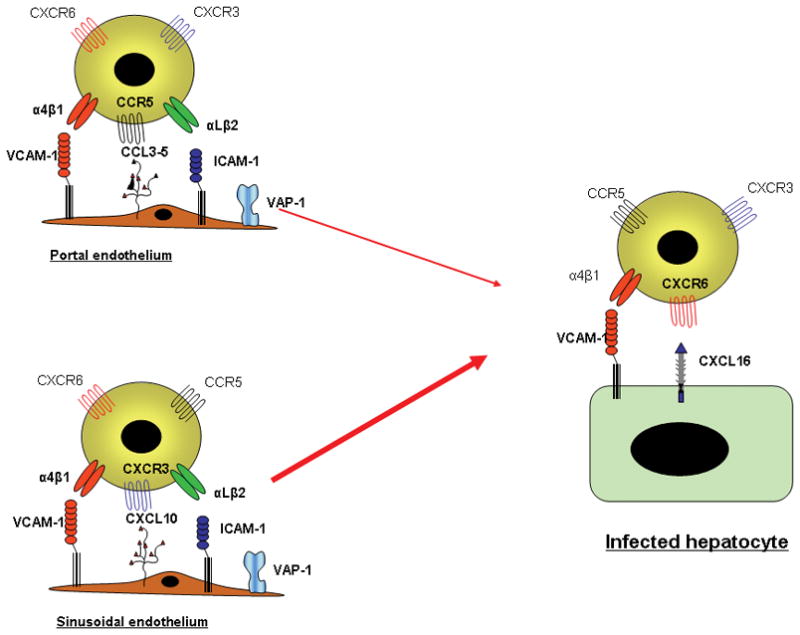
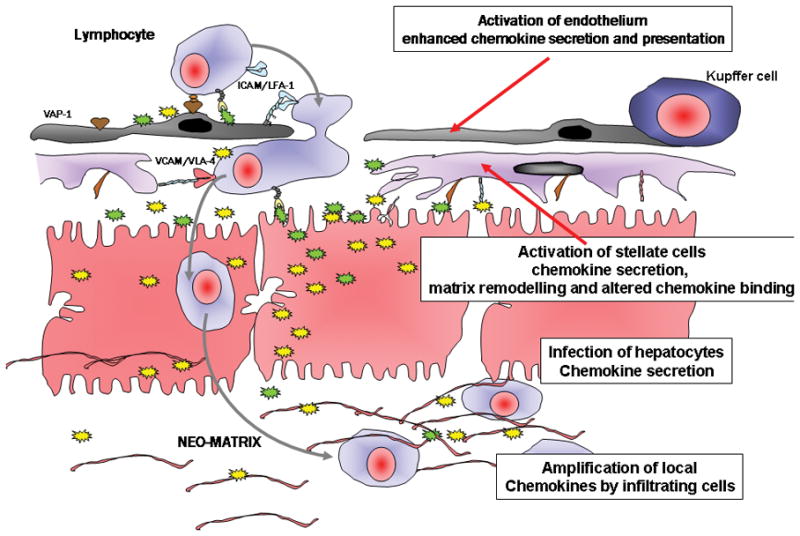
Chronic infection with the hepatitis C virus, a noncytopathic hepatotropic RNA virus, affects over 170 million people worldwide. In the majority of cases, neither the early innate immune response nor the later adaptive immune response succeeds in clearing the virus, and the infection becomes chronic. Furthermore, in many patients, the ineffective inflammatory response drives fibrogenesis and the development of cirrhosis. It is critical to understand this immune pathology if preventative and curative therapies are to be developed. Chemokines are a superfamily of small proteins that promote leukocyte migration and orchestrate the immune response to viruses, including hepatitis C virus. Chemokines are crucial for viral elimination, but inappropriate persistence of expression in chronic hepatitis C infection can drive tissue damage and inflammation. Here we review the role of chemokines and their receptors in hepatitis C virus infection.
Figures
References
-
- Lauer GM, Walker BD. Hepatitis C virus infection. N Engl J Med. 2001;345:41–52. - PubMed
-
- Rehermann B, Nascimbeni M. Immunology of hepatitis B virus and hepatitis C virus infection. Nat Rev Immunol. 2005;5:215–229. - PubMed
-
- Dustin LB, Rice CM. Flying under the radar: the immunobiology of hepatitis C. Annu Rev Immunol. 2007;25:71–99. - PubMed
-
- Rot A, von Andrian UH. Chemokines in innate and adaptive host defense: basic chemokinese grammar for immune cells. Annu Rev Immunol. 2004;22:891–928. - PubMed
-
- Viola A, Luster AD. Chemokines and their receptors: drug targets in immunity and inflammation. Annu Rev Pharmacol Toxicol. 2008;48:171–197. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical