

Tanycyte pyroglutamyl peptidase II contributes to regulation of the hypothalamic-pituitary-thyroid axis through glial-axonal associations in the median eminence
- PMID: 19179432
- PMCID: PMC2671897
- DOI: 10.1210/en.2008-1643
Tanycyte pyroglutamyl peptidase II contributes to regulation of the hypothalamic-pituitary-thyroid axis through glial-axonal associations in the median eminence
Abstract
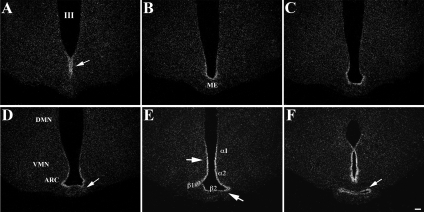
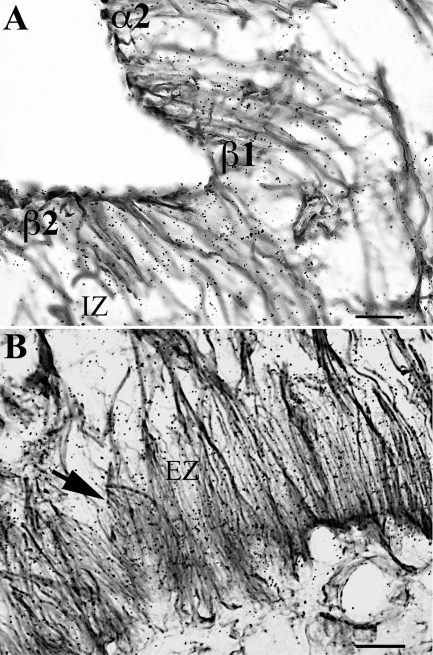
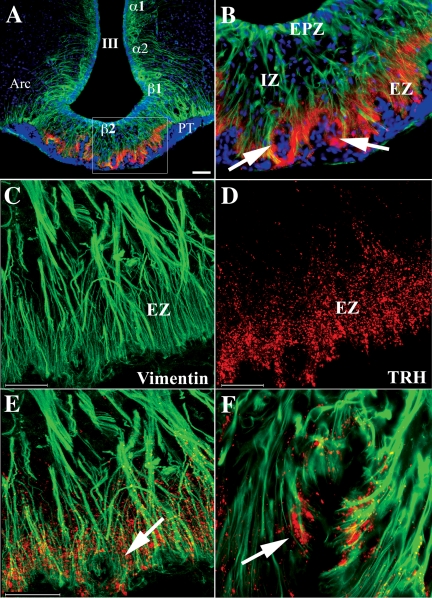
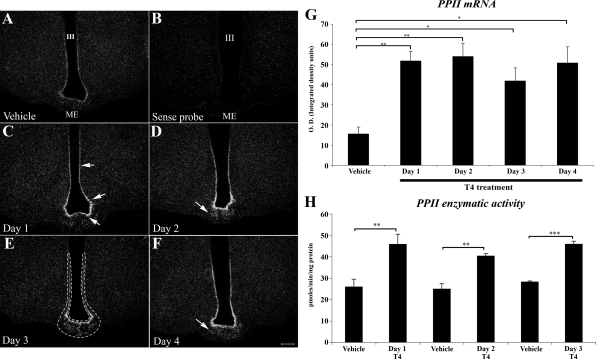
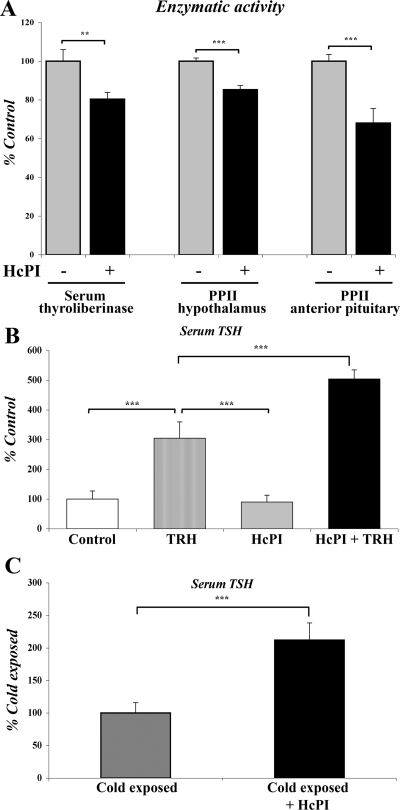
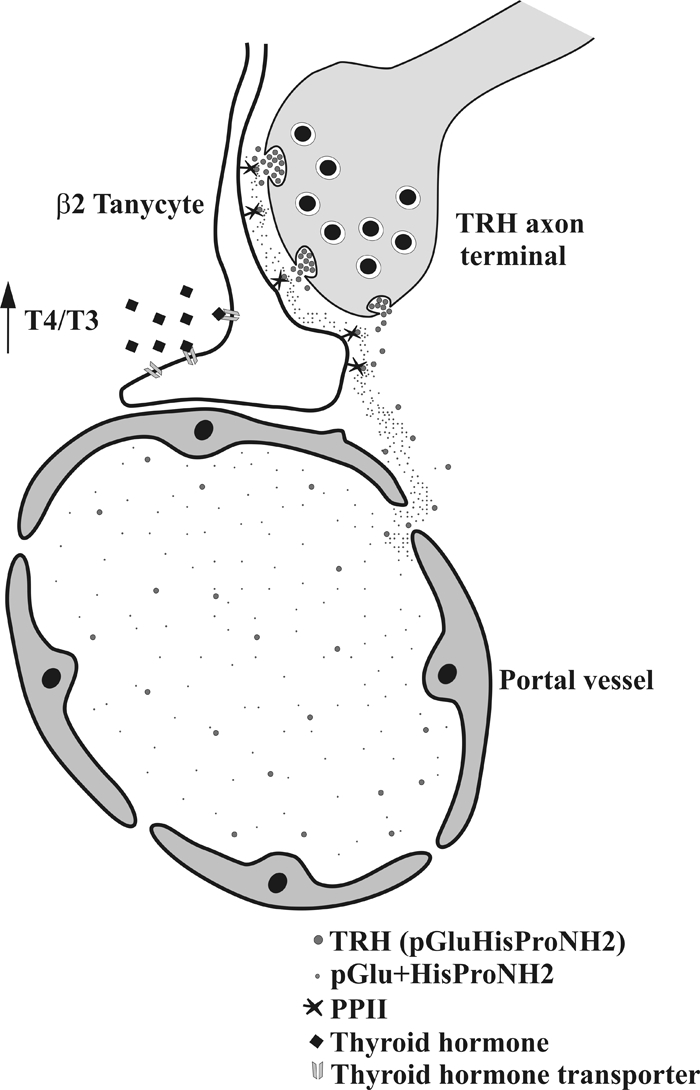
Pyroglutamyl peptidase II (PPII), a highly specific membrane-bound metallopeptidase that inactivates TRH in the extracellular space, is tightly regulated by thyroid hormone in cells of the anterior pituitary. Whether PPII has any role in the region where axons containing hypophysiotropic TRH terminate, the median eminence, is unknown. For this purpose, we analyzed the cellular localization and regulation of PPII mRNA in the mediobasal hypothalamus in adult, male rats. PPII mRNA was localized in cells lining the floor and infralateral walls of the third ventricle and coexpressed with vimentin, establishing these cells as tanycytes. PPII mRNA extended in a linear fashion from the tanycyte cell bodies in the base of the third ventricle to its cytoplasmic and end-feet processes in the external zone of the median eminence in close apposition to pro-TRH-containing axon terminals. Compared with vehicle-treated, euthyroid controls, animals made thyrotoxic by the i.p. administration of 10 microg L-T(4) daily for 1-3 d, showed dramatically increased accumulation of silver grains in the mediobasal hypothalamus and an approximately 80% increase in enzymatic activity. PPII inhibition in mediobasal hypothalamic explants increased TRH secretion, whereas i.p. injection of a specific PPII inhibitor increased cold stress- and TRH-induced TSH levels in plasma. We propose that an increase in circulating thyroid hormone up-regulates PPII activity in tanycytes and enhances degradation of extracellular TRH in the median eminence through glial-axonal associations, contributing to the feedback regulation of thyroid hormone on anterior pituitary TSH secretion.
Figures
Comment in
-
The ups and downs of thyrotropin-releasing hormone.Endocrinology. 2009 May;150(5):2021-3. doi: 10.1210/en.2009-0261. Endocrinology. 2009. PMID: 19383877 Free PMC article. No abstract available.
Similar articles
-
Fasting Enhances Pyroglutamyl Peptidase II Activity in Tanycytes of the Mediobasal Hypothalamus of Male Adult Rats.Endocrinology. 2015 Jul;156(7):2713-23. doi: 10.1210/en.2014-1885. Epub 2015 May 5. Endocrinology. 2015. PMID: 25942072
-
Thyroxine-induced expression of pyroglutamyl peptidase II and inhibition of TSH release precedes suppression of TRH mRNA and requires type 2 deiodinase.J Endocrinol. 2011 Oct;211(1):73-8. doi: 10.1530/JOE-11-0248. Epub 2011 Jul 25. J Endocrinol. 2011. PMID: 21788297 Free PMC article.
-
Mediobasal hypothalamic and adenohypophyseal TRH-degrading enzyme (PPII) is down-regulated by zinc deficiency.Int J Dev Neurosci. 2015 Nov;46:115-24. doi: 10.1016/j.ijdevneu.2015.08.001. Epub 2015 Aug 24. Int J Dev Neurosci. 2015. PMID: 26315400
-
TRH inactivation in the extracellular compartment: role of pyroglutamyl peptidase II.Neurobiology (Bp). 1998;6(1):45-57. Neurobiology (Bp). 1998. PMID: 9713831 Review.
-
Tanycytes and the Control of Thyrotropin-Releasing Hormone Flux Into Portal Capillaries.Front Endocrinol (Lausanne). 2019 Jun 25;10:401. doi: 10.3389/fendo.2019.00401. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31293518 Free PMC article. Review.
Cited by
-
The influence of extended fasting on thyroid hormone: local and differentiated regulatory mechanisms.Front Endocrinol (Lausanne). 2024 Aug 26;15:1443051. doi: 10.3389/fendo.2024.1443051. eCollection 2024. Front Endocrinol (Lausanne). 2024. PMID: 39253586 Free PMC article. Review.
-
Understanding the hypothalamus-pituitary-thyroid axis in mct8 deficiency.Eur Thyroid J. 2012 Jul;1(2):72-9. doi: 10.1159/000339474. Epub 2012 Jun 20. Eur Thyroid J. 2012. PMID: 24783000 Free PMC article.
-
Advances in TRH signaling.Rev Endocr Metab Disord. 2016 Dec;17(4):545-558. doi: 10.1007/s11154-016-9375-y. Rev Endocr Metab Disord. 2016. PMID: 27515033 Review.
-
Tanycytes control the hormonal output of the hypothalamic-pituitary-thyroid axis.Nat Commun. 2017 Sep 7;8(1):484. doi: 10.1038/s41467-017-00604-6. Nat Commun. 2017. PMID: 28883467 Free PMC article.
-
Hypothalamic tanycytes: potential roles in the control of feeding and energy balance.Trends Neurosci. 2013 Feb;36(2):91-100. doi: 10.1016/j.tins.2012.12.008. Epub 2013 Jan 17. Trends Neurosci. 2013. PMID: 23332797 Free PMC article. Review.
References
-
- Charli JL, Vargas MA, Cisneros M, de Gortari P, Baeza MA, Jasso P, Bourdais J, Perez L, Uribe RM, Joseph-Bravo P 1998 TRH inactivation in the extracellular compartment: role of pyroglutamyl peptidase II. Neurobiology 6:45–57 - PubMed
-
- Vargas M, Mendez M, Cisneros M, Joseph-Bravo P, Charli JL 1987 Regional distribution of the membrane-bound pyroglutamate amino peptidase-degrading thyrotropin-releasing hormone in rat brain. Neurosci Lett 79:311–314 - PubMed
-
- Heuer H, Schäfer MK, O'Donnell D, Walker P, Bauer K 2000 Expression of thyrotropin-releasing hormone receptor 2 (TRH-R2) in the central nervous system of rats. J Comp Neurol 428:319–336 - PubMed
-
- Schmitmeier S, Thole H, Bader A, Bauer K 2002 Purification and characterization of the thyrotropin-releasing hormone (TRH)-degrading serum enzyme and its identification as a product of liver origin. Eur J Biochem 269:1278–1286 - PubMed
-
- Grosvenor CE, Mena F 1980 Evidence that thyrotropin-releasing hormone and a hypothalamic prolactin-releasing factor may function in the release of prolactin in the lactating rat. Endocrinology 107:863–868 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
