

Both thiamine uptake and biosynthesis of thiamine precursors are required for intracellular replication of Listeria monocytogenes
- PMID: 19181806
- PMCID: PMC2655527
- DOI: 10.1128/JB.01636-08
Both thiamine uptake and biosynthesis of thiamine precursors are required for intracellular replication of Listeria monocytogenes
Abstract
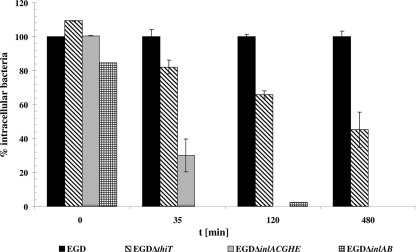
Thiamine pyrophosphate is an essential cofactor involved in central metabolism and amino acid biosynthesis and is derived from thiamine (vitamin B(1)). The extent to which this metabolite is available to bacterial pathogens replicating within host cells is still little understood. Growth studies using modified minimal Welshimer's broth (mMWB) supplemented with thiamine or the thiamine precursor hydroxymethylpyrimidine (HMP) showed that Listeria monocytogenes, in agreement with bioinformatic prediction, is able to synthesize thiamine only in the presence of HMP. This appears to be due to a lack of ThiC, which is involved in HMP synthesis. The knockout of thiD (lmo0317), which probably catalyzes the phosphorylation of HMP, inhibited growth in mMWB supplemented with HMP and reduced the replication rate of L. monocytogenes in epithelial cells. Mutation of a predicted thiamine transporter gene, lmo1429, led to reduced proliferation of L. monocytogenes in mMWB containing thiamine or thiamine phosphates and also within epithelial cells but had no influence on the expression of the virulence factors Hly and ActA. The toxic thiamine analogue pyrithiamine inhibited growth of wild-type strain EGD but not of the transporter mutant EGDDeltathiT. We also demonstrated that ThiT binds thiamine, a finding compatible with ThiT acting as the substrate-binding component of a multimeric thiamine transporter complex. These data provide experimental evidence that Lmo1429 homologs including Bacillus YuaJ are necessary for thiamine transport in gram-positive bacteria and are therefore proposed to be annotated "ThiT." Taken together, these data indicate that concurrent thiamine uptake and biosynthesis of thiamine precursors is a strategy of L. monocytogenes and possibly other facultative intracellular pathogens to enable proliferation within the cytoplasm.
Figures






Similar articles
-
Generation of branched-chain fatty acids through lipoate-dependent metabolism facilitates intracellular growth of Listeria monocytogenes.J Bacteriol. 2009 Apr;191(7):2187-96. doi: 10.1128/JB.01179-08. Epub 2009 Jan 30. J Bacteriol. 2009. PMID: 19181817 Free PMC article.
-
Thiamine plays a critical role in the acid tolerance of Listeria monocytogenes.FEMS Microbiol Lett. 2012 Jan;326(2):137-43. doi: 10.1111/j.1574-6968.2011.02442.x. Epub 2011 Nov 14. FEMS Microbiol Lett. 2012. PMID: 22092299
-
Thioredoxin A Is Essential for Motility and Contributes to Host Infection of Listeria monocytogenes via Redox Interactions.Front Cell Infect Microbiol. 2017 Jun 28;7:287. doi: 10.3389/fcimb.2017.00287. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28702378 Free PMC article.
-
Metabolism of the Gram-Positive Bacterial Pathogen Listeria monocytogenes.Microbiol Spectr. 2019 Jul;7(4):10.1128/microbiolspec.gpp3-0066-2019. doi: 10.1128/microbiolspec.GPP3-0066-2019. Microbiol Spectr. 2019. PMID: 31418407 Free PMC article. Review.
-
ActA of Listeria monocytogenes and Its Manifold Activities as an Important Listerial Virulence Factor.Curr Top Microbiol Immunol. 2017;399:113-132. doi: 10.1007/82_2016_30. Curr Top Microbiol Immunol. 2017. PMID: 27726006 Review.
Cited by
-
Two essential arginine residues in the T components of energy-coupling factor transporters.J Bacteriol. 2009 Nov;191(21):6482-8. doi: 10.1128/JB.00965-09. Epub 2009 Aug 28. J Bacteriol. 2009. PMID: 19717603 Free PMC article.
-
Wholly Rickettsia! Reconstructed Metabolic Profile of the Quintessential Bacterial Parasite of Eukaryotic Cells.mBio. 2017 Sep 26;8(5):e00859-17. doi: 10.1128/mBio.00859-17. mBio. 2017. PMID: 28951473 Free PMC article.
-
Bacterial leaf blight resistance in rice: a review of conventional breeding to molecular approach.Mol Biol Rep. 2019 Feb;46(1):1519-1532. doi: 10.1007/s11033-019-04584-2. Epub 2019 Jan 9. Mol Biol Rep. 2019. PMID: 30628024 Review.
-
The structural basis of modularity in ECF-type ABC transporters.Nat Struct Mol Biol. 2011 Jun 26;18(7):755-60. doi: 10.1038/nsmb.2073. Nat Struct Mol Biol. 2011. PMID: 21706007
-
Comparative Genomic Analysis of Fusobacterium necrophorum Provides Insights into Conserved Virulence Genes.Microbiol Spectr. 2022 Dec 21;10(6):e0029722. doi: 10.1128/spectrum.00297-22. Epub 2022 Oct 7. Microbiol Spectr. 2022. PMID: 36219094 Free PMC article.
References
-
- Becker, D., M. Selbach, C. Rollenhagen, M. Ballmaier, T. F. Meyer, M. Mann, and D. Bumann. 2006. Robust Salmonella metabolism limits possibilities for new antimicrobials. Nature 440303-307. - PubMed
-
- Begley, T. P., D. M. Downs, S. E. Ealick, F. W. McLafferty, A. P. Van Loon, S. Taylor, N. Campobasso, H. J. Chiu, C. Kinsland, J. J. Reddick, and J. Xi. 1999. Thiamin biosynthesis in prokaryotes. Arch. Microbiol. 171293-300. - PubMed
-
- Bergmann, B., D. Raffelsbauer, M. Kuhn, M. Goetz, S. Hom, and W. Goebel. 2002. InlA- but not InlB-mediated internalization of Listeria monocytogenes by non-phagocytic mammalian cells needs the support of other internalins. Mol. Microbiol. 43557-570. - PubMed
-
- Bettendorff, L. 1995. Thiamine homeostasis in neuroblastoma cells. Neurochem. Int. 26295-302. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
