

Tumor initiating cells in malignant gliomas: biology and implications for therapy
- PMID: 19189072
- PMCID: PMC2693383
- DOI: 10.1007/s00109-009-0440-9
Tumor initiating cells in malignant gliomas: biology and implications for therapy
Abstract
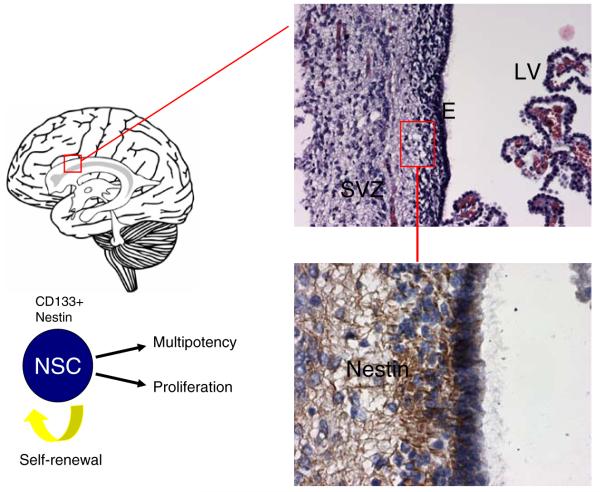
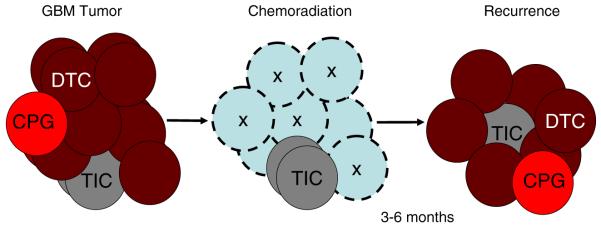
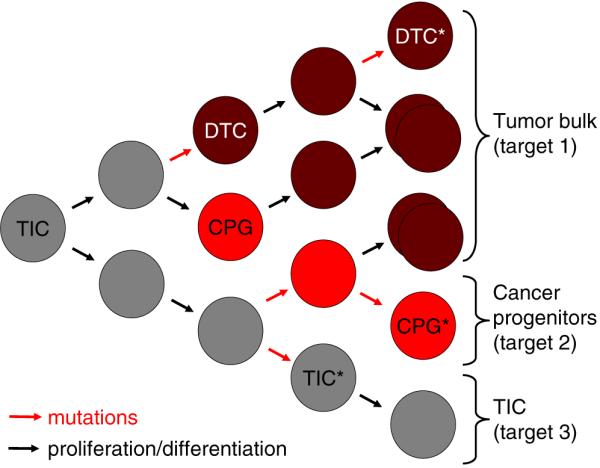
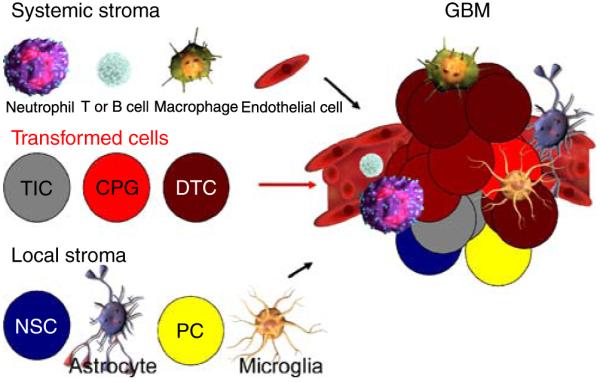
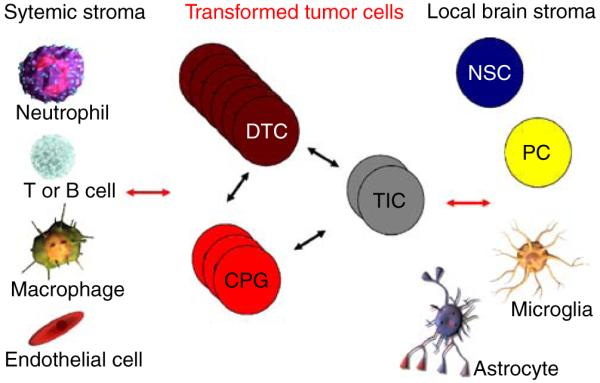
A rare subpopulation of cells within malignant gliomas, which shares canonical properties with neural stem cells (NSCs), may be integral to glial tumor development and perpetuation. These cells, also known as tumor initiating cells (TICs), have the ability to self-renew, develop into any cell in the overall tumor population (multipotency), and proliferate. A defining property of TICs is their ability to initiate new tumors in immunocompromised mice with high efficiency. Mounting evidence suggests that TICs originate from the transformation of NSCs and their progenitors. New findings show that TICs may be more resistant to chemotherapy and radiation than the bulk of tumor cells, thereby permitting recurrent tumor formation and accounting for the failure of conventional therapies. The development of new therapeutic strategies selectively targeting TICs while sparing NSCs may provide for more effective treatment of malignant gliomas.
Figures

References
-
- Jordan CT, Guzman ML, Noble M. Cancer stem cells. N Engl J Med. 2006;355:1253–1261. - PubMed
-
- Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. - PubMed
-
- Al-Hajj M, Becker MW, Wicha M, Weissman I, Clarke MF. Therapeutic implications of cancer stem cells. Curr Opin Genet Dev. 2004;14:43–47. - PubMed
-
- Wicha MS, Liu S, Dontu G. Cancer stem cells: an old idea—a paradigm shift. Cancer Res. 2006;66:1883–1890. discussion 1895-1886. - PubMed
-
- Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, Minden M, Paterson B, Caligiuri MA, Dick JE. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature. 1994;367:645–648. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
