

doi: 10.1093/nar/gkp050.
Epub 2009 Feb 3.
U1-independent pre-mRNA splicing contributes to the regulation of alternative splicing
Affiliations
- PMID: 19190090
- PMCID: PMC2665233
- DOI: 10.1093/nar/gkp050
Item in Clipboard
U1-independent pre-mRNA splicing contributes to the regulation of alternative splicing
Nucleic Acids Res.
2009 Apr.
Abstract
U1 snRNP plays a crucial role in the 5' splice site recognition during splicing. Here we report the first example of naturally occurring U1-independent U2-type splicing in humans. The U1 components were not included in the pre-spliceosomal E complex formed on the human F1gamma (hF1gamma) intron 9 in vitro. Moreover, hF1gamma intron 9 was efficiently spliced even in U1-disrupted Xenopus oocytes as well as in U1-inactivated HeLa nuclear extracts. Finally, hF1gamma exon 9 skipping induced by an alternative splicing regulator Fox-1 was impaired when intron 9 was changed to the U1-dependent one. Our results suggest that U1-independent splicing contributes to the regulation of alternative splicing of a class of pre-mRNAs.
Figures
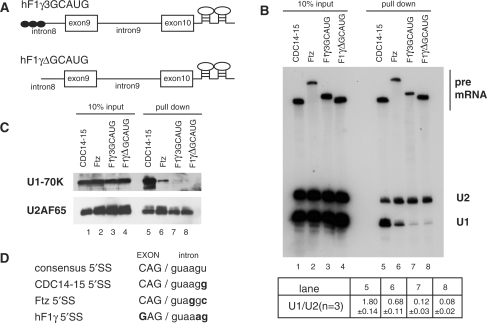
Purification of the pre-spliceosomal E complex formed on hF1γ exon 9–10 pre-mRNA in vitro. (A) The schematic representation of pre-mRNAs, CDC14-15, ftz, hF1γ3GCAUG and hF1γΔGCAUG, fused with two MS2-binding sites at the 3′ exons. Boxes and lines represent exons and introns, respectively. The Fox-1 binding element GCAUG is shown as a closed circle. (B) Northern blotting of the purified E complexes with U1 and U2 snRNA probes (lanes 5–8) and aliquots of the reaction mixtures (lanes 1–4). Average and standard deviation (SD) of the ratio of U1 snRNA to U2 snRNA from three independent experiments are shown at the bottom. (C) Western blotting of the purified E complexes on the pre-mRNAs using U1-70K and U2AF65 antibodies (lanes 5–8) and aliquots of the reaction mixtures (lanes 1–4). (D) The sequences of 5′ splice sites of four kinds of pre-mRNAs as shown in (A). Capital and lowercase letters correspond to exonic and intronic residues, respectively. The nucleotide different from the consensus site is shown in bold.
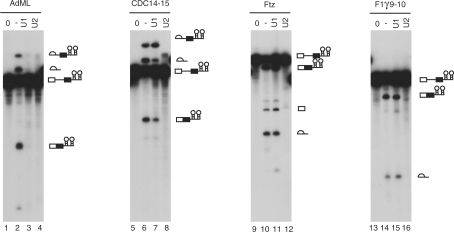
hF1γ exon 9–10 pre-mRNA is spliced efficiently in U1-inactivated HeLa nuclear extracts in vitro. Splicing reaction of AdML (lanes 1–4), CDC14-15 (lanes 5–8), ftz (lanes 9–12) and hF1γ 9–10 (lanes 13–16) with the normal HeLa cell nuclear extracts (lanes 2, 6, 10 and 14), the U1- (lanes 3, 7, 11 and 15) or U2-inactivated (lanes 4, 8, 12 and 16) extracts, as well as transcripts without incubation (lanes 1, 5, 9 and 13). Pre-mRNA, intermediates and splicing products are indicated schematically on the right.
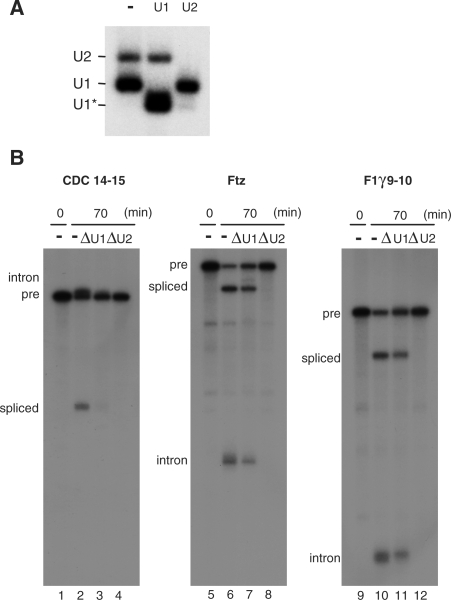
Splicing of hF1γ exon 9–10 pre-mRNA in U1- or U2-disrupted Xenopus oocytes. (A) Northern blotting of endogenous U1 and U2 snRNAs in Xenopus oocytes. Antisense oligonucleotide against U1 or U2 snRNA was injected into the cytoplasm of Xenopus oocytes to disrupt endogenous U1 or U2. The same volume of water was injected as a control (–). Note that U1 snRNA was truncated by the injection of the U1 antisense oligo (the fast migrating band indicated as U1*), since the oligo targets the 5′ end of U1 snRNA. (B) Splicing of the 32P-labeled CDC14-15, ftz and hF1γ 9–10 pre-mRNAs in the nuclei of control (lanes 2, 6 and 10), U1-disrupted (lanes 3, 7 and 11), or U2-disrupted oocytes (lanes 4, 8 and 12). The hF1γ 9–10 transcript lacks the intron 8 sequence. RNA was analyzed immediately (lanes 1, 5 and 9) or at 70 min (lanes 2–4, 6–8 and 10–12) after injection.
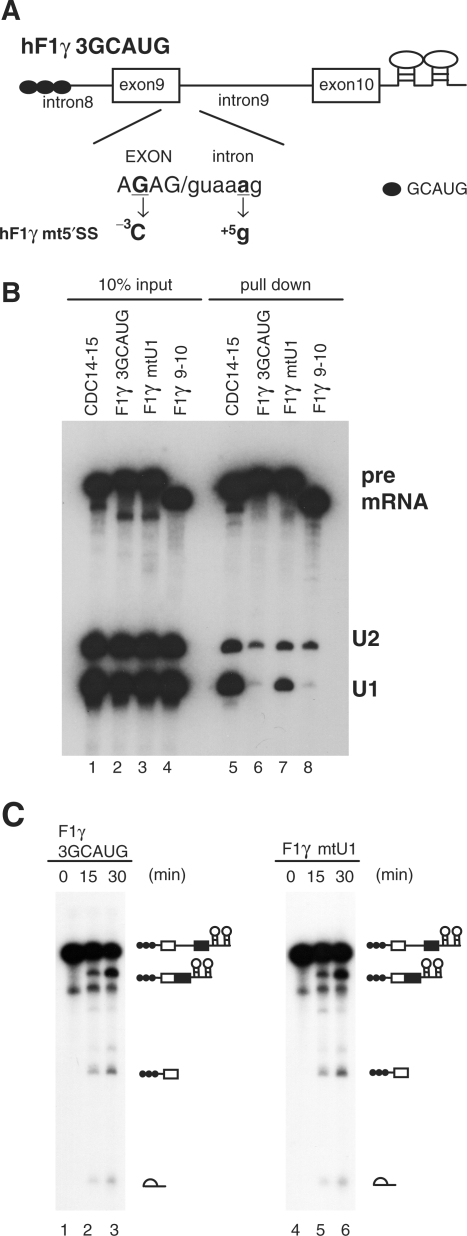
Role of the 5′ splice site sequence in U1-independent splicing. (A) Base-substitutions introduced into the 5′ splice site of hF1γ3GCAUG pre-mRNA are shown (–3G > C and +5a > g). Capital and lowercase letters correspond to exonic and intronic nucleotides, respectively. Boxes and lines represent exons and introns, respectively. The Fox-1 binding element GCAUG is shown as a closed circle. (B) Northern blotting of the purified E complexes with U1 and U2 snRNA probes (lanes 5–8) and aliquots of the reaction mixtures (lanes 1–4). (C) Splicing reaction of hF1γ3GCAUG (lanes 1–3) and hF1γmt5′SS (lanes 4–6). These pre-mRNAs were incubated under the standard in vitro splicing condition for indicated time above each lane. Pre-mRNA, intermediates and splicing products are indicated schematically on the right.
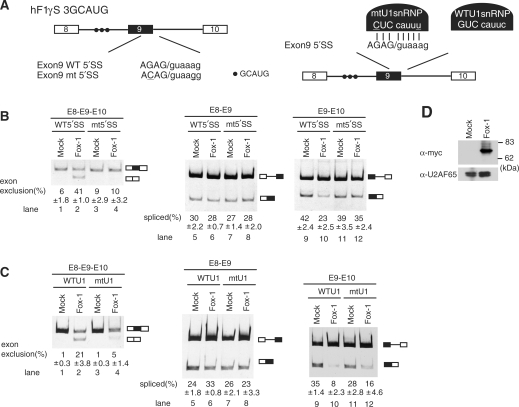
U1-independent splicing is indispensable for the regulation of alternative splicing of hF1γ by Fox-1. (A) The schematic representation of wild-type (WT) and mutant (mtU1) hF1γ mini-genes (left) and the suppressor U1 snRNP (right). The mutations introduced into the 5′ splice site of exon 9 are underlined. Closed circles indicate the Fox-1 binding element. (B) Transfection assay of hF1γ WT and 5′SSmt mini-genes co-expressed with pCS2+MT vector or pCS+MT-Fox-1 in HeLa cells. The left panel shows spliced products amplified by the oligonucleotides annealed with exons 8 and 10. The middle and right panels show the splicing reaction between exons 8 and 9 and between exons 9 and 10. Positions of splicing products and unspliced transcripts are schematically shown on the right. All experiments were independently performed four times. Average and standard deviation of exon 9 exclusion and splicing efficiency are shown at the bottom of each lane. (C) Suppressor U1 snRNA experiments in HeLa cells. The wild-type or mutant U1 snRNA was co-expressed with wild-type hF1γ mini-gene and pCS2+MT vector or pCS+MT-Fox-1. The spliced products were analyzed by RT-PCR using the oligonucleotides annealed with exons 8 and 10 (left panel), exons 8 and 9 (middle panel) and exons 9 and 10 (right panel). Positions of splicing products and unspliced transcripts are schematically shown on the right. All experiments were independently performed three times. Average and standard deviation of exon 9 exclusion and splicing efficiency are shown at the bottom of each lane. (D) Western blotting of cell extracts using the anti-Myc antibody (upper panel) and anti-U2AF65 antibody as a loading control (lower panel). Molecular size markers are shown on the right.
Similar articles
-
Tissue-specific splicing regulator Fox-1 induces exon skipping by interfering E complex formation on the downstream intron of human F1gamma gene.Nucleic Acids Res. 2007;35(16):5303-11. doi: 10.1093/nar/gkm569. Epub 2007 Aug 7. Nucleic Acids Res. 2007. PMID: 17686786 Free PMC article.
-
Genome-wide RNA-binding analysis of the trypanosome U1 snRNP proteins U1C and U1-70K reveals cis/trans-spliceosomal network.Nucleic Acids Res. 2014 Jun;42(10):6603-15. doi: 10.1093/nar/gku286. Epub 2014 Apr 19. Nucleic Acids Res. 2014. PMID: 24748659 Free PMC article.
-
The transition in spliceosome assembly from complex E to complex A purges surplus U1 snRNPs from alternative splice sites.Nucleic Acids Res. 2012 Aug;40(14):6850-62. doi: 10.1093/nar/gks322. Epub 2012 Apr 13. Nucleic Acids Res. 2012. PMID: 22505580 Free PMC article.
-
Principles and correction of 5'-splice site selection.RNA Biol. 2022 Jan;19(1):943-960. doi: 10.1080/15476286.2022.2100971. RNA Biol. 2022. PMID: 35866748 Free PMC article. Review.
-
Role and mechanism of U1-independent pre-mRNA splicing in the regulation of alternative splicing.RNA Biol. 2009 Sep-Oct;6(4):395-8. doi: 10.4161/rna.6.4.9318. Epub 2009 Sep 21. RNA Biol. 2009. PMID: 19667760 Review.
Cited by
-
U1 snRNP Telescripting: Suppression of Premature Transcription Termination in Introns as a New Layer of Gene Regulation.Cold Spring Harb Perspect Biol. 2019 Feb 1;11(2):a032235. doi: 10.1101/cshperspect.a032235. Cold Spring Harb Perspect Biol. 2019. PMID: 30709878 Free PMC article. Review.
-
U1 snRNA interactions with deep intronic sequences regulate splicing of multiple exons of spinal muscular atrophy genes.Front Neurosci. 2024 Jul 12;18:1412893. doi: 10.3389/fnins.2024.1412893. eCollection 2024. Front Neurosci. 2024. PMID: 39086841 Free PMC article.
-
Pick one, but be quick: 5' splice sites and the problems of too many choices.Genes Dev. 2013 Jan 15;27(2):129-44. doi: 10.1101/gad.209759.112. Genes Dev. 2013. PMID: 23348838 Free PMC article. Review.
-
Dramatically reduced spliceosome in Cyanidioschyzon merolae.Proc Natl Acad Sci U S A. 2015 Mar 17;112(11):E1191-200. doi: 10.1073/pnas.1416879112. Epub 2015 Mar 2. Proc Natl Acad Sci U S A. 2015. PMID: 25733880 Free PMC article.
-
Mechanisms of activation and repression by the alternative splicing factors RBFOX1/2.RNA. 2012 Feb;18(2):274-83. doi: 10.1261/rna.030486.111. Epub 2011 Dec 19. RNA. 2012. PMID: 22184459 Free PMC article.
References
-
- Graveley BR. Alternative splicing: increasing diversity in the proteomic world. Trends Genet. 2001;17:100–107. - PubMed
-
- Maniatis T, Tasic B. Alternative pre-mRNA splicing and proteome expansion in metazoans. Nature. 2002;418:236–243. - PubMed
-
- Black DL. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003;72:291–336. - PubMed
-
- Ben-Dov C, Hartmann B, Lundgren J, Valcarcel J. Genome-wide analysis of alternative pre-mRNA splicing. J. Biol. Chem. 2008;283:1229–1233. - PubMed
-
- Das R, Zhou Z, Reed R. Functional association of U2 snRNP with the ATP-independent spliceosomal complex E. Mol. Cell. 2000;5:779–787. - PubMed
