

Galectin-3 mediates nuclear beta-catenin accumulation and Wnt signaling in human colon cancer cells by regulation of glycogen synthase kinase-3beta activity
- PMID: 19190323
- PMCID: PMC2990400
- DOI: 10.1158/0008-5472.CAN-08-4153
Galectin-3 mediates nuclear beta-catenin accumulation and Wnt signaling in human colon cancer cells by regulation of glycogen synthase kinase-3beta activity
Abstract
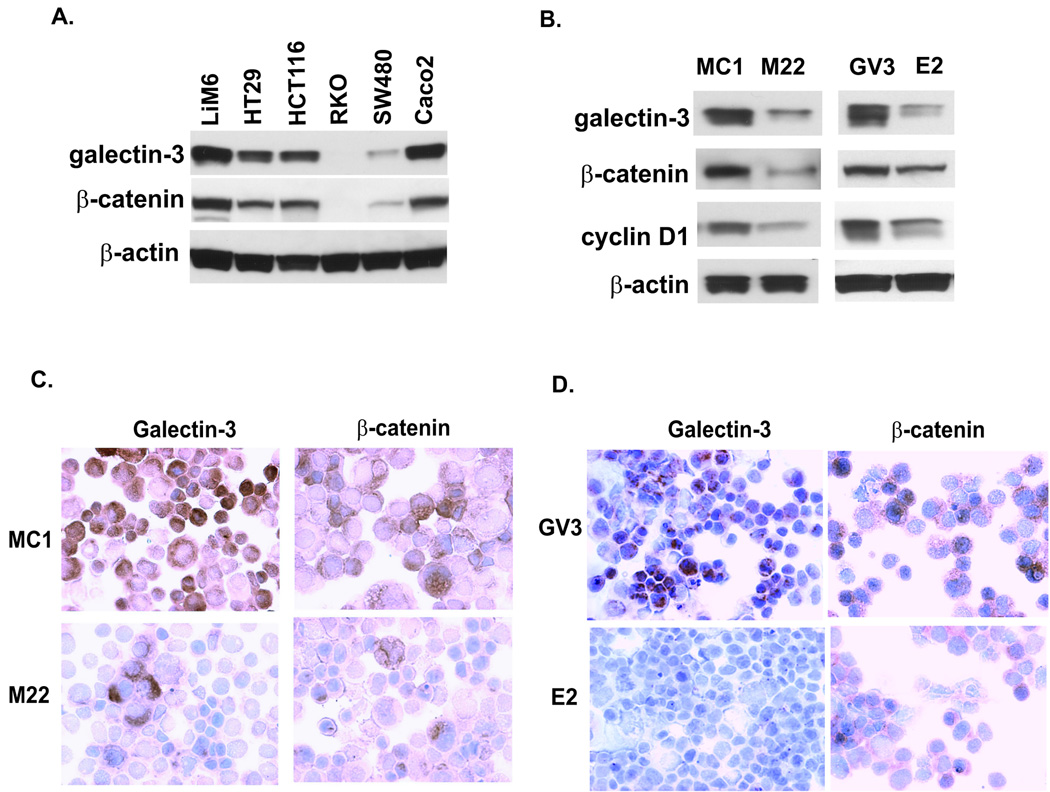
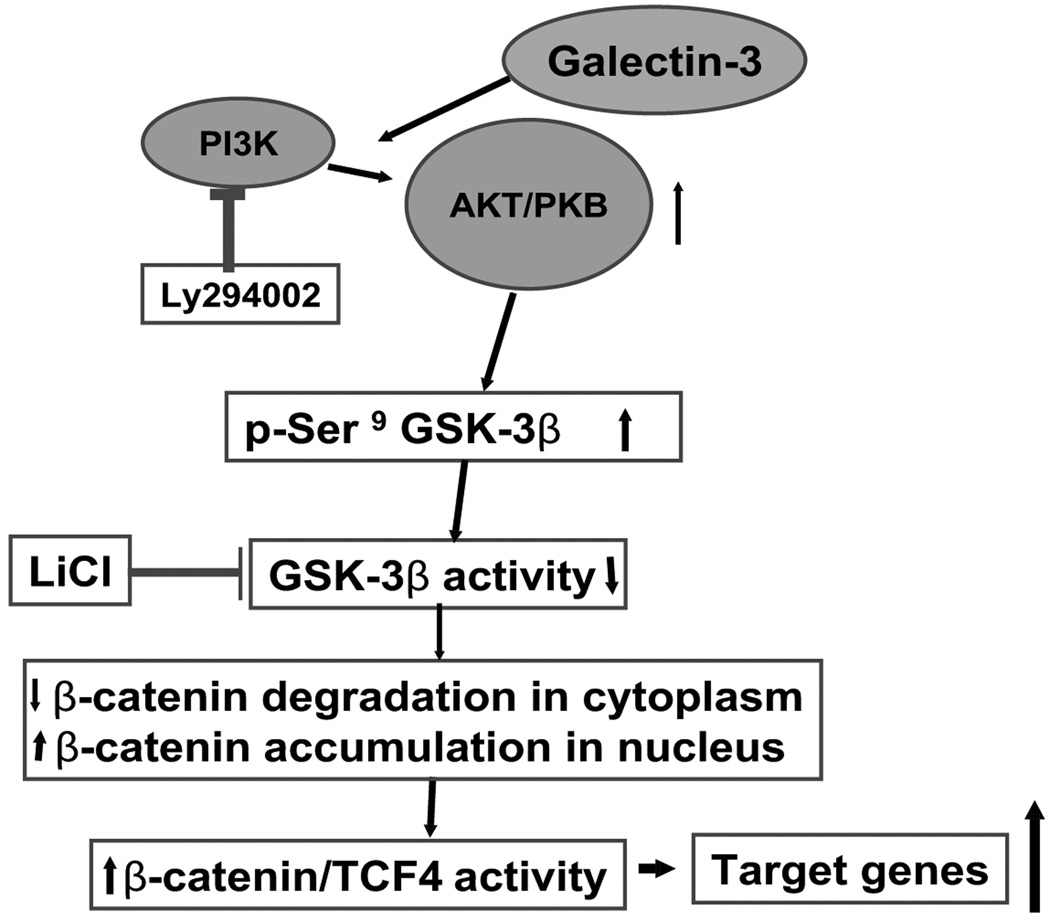
Wnt/beta-catenin signaling plays an essential role in colon carcinogenesis. Galectin-3, a beta-galactoside-binding protein, has been implicated in Wnt signaling, but the precise mechanisms by which galectin-3 modulates the Wnt pathway are unknown. In the present study, we determined the effects of galectin-3 on the Wnt/beta-catenin pathway in colon cancer cells, as well as the mechanisms involved. Galectin-3 levels were manipulated in human colon cancer cells by stable transfection of galectin-3 antisense, short hairpin RNA, or full-length galectin-3 cDNA, and effects on beta-catenin levels, subcellular distribution, and Wnt signaling were determined. Galectin-3 levels correlated with beta-catenin levels in a variety of colon cancer cell lines. Down-regulation of galectin-3 resulted in decreased beta-catenin protein levels but no change in beta-catenin mRNA levels, suggesting that galectin-3 modulates beta-catenin by another mechanism. Reduction of galectin-3 led to reduced nuclear beta-catenin with a concomitant decrease in TCF4 transcriptional activity and expression of its target genes. Conversely, transfection of galectin-3 cDNA into colon cancer cells increased beta-catenin expression and TCF4 transcriptional activity. Down-regulation of galectin-3 resulted in AKT and glycogen synthase kinase-3beta (GSK-3beta) dephosphorylation and increased GSK activity, increasing beta-catenin phosphorylation and degradation. Ly294002, an inhibitor of phosphatidylinositol 3-kinase, and dominant-negative AKT, suppressed TCF4 transcriptional activity induced by galectin-3 whereas LiCl, a GSK-3beta inhibitor, increased TCF4 activity, mimicking the effects of galectin-3. These results suggest that galectin-3 mediates Wnt signaling, at least in part, by regulating GSK-3beta phosphorylation and activity via the phosphatidylinositol 3-kinase/AKT pathway, and, thus, the degradation of beta-catenin in colon cancer cells.
Figures




References
-
- Kolligs FT, Bommer G, Goke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion. 2002;66:131–144. - PubMed
-
- Giles RH, van Es JH, Clevers H. Caught up in a Wnt storm: Wnt signaling in cancer. Biochim Biophys Acta. 2003;1653:1–24. - PubMed
-
- Behrens J, Jerchow BA, Wurtele M, Grimm J, Asbrand C, Wirtz R, Kuhl M, Wedlich D, Birchmeier W. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science. 1998;280:596–599. - PubMed
-
- Sparks AB, Morin PJ, Vogelstein B, Kinzler KW. Mutational analysis of the APC/beta-catenin/Tcf pathway in colorectal cancer. Cancer Res. 1998;58:1130–1134. - PubMed
-
- Rubinfeld B, Albert I, Porfiri E, Fiol C, Munemitsu S, Polakis P. Binding of GSK3beta to the APC-beta-catenin complex and regulation of complex assembly. Science. 1996;272:1023–1026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
