

Sequential metabolism of 7-dehydrocholesterol to steroidal 5,7-dienes in adrenal glands and its biological implication in the skin
- PMID: 19190754
- PMCID: PMC2629546
- DOI: 10.1371/journal.pone.0004309
Sequential metabolism of 7-dehydrocholesterol to steroidal 5,7-dienes in adrenal glands and its biological implication in the skin
Abstract
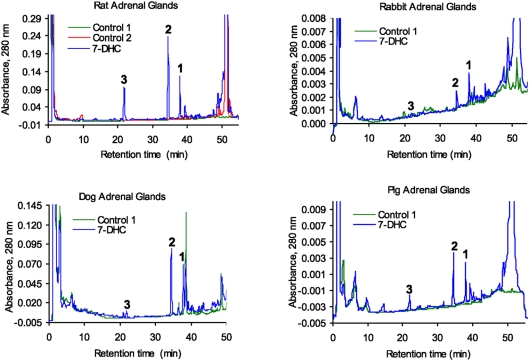
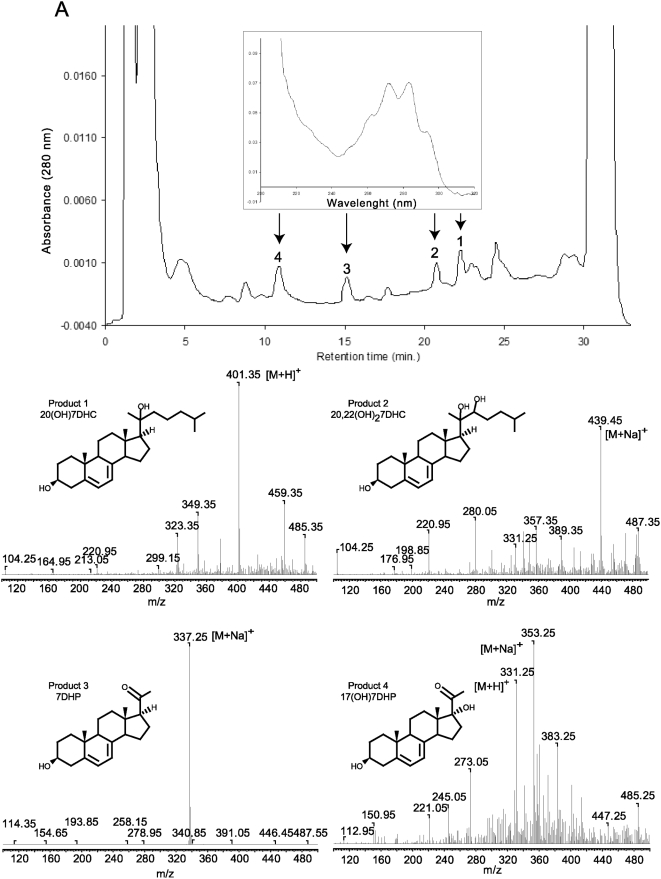
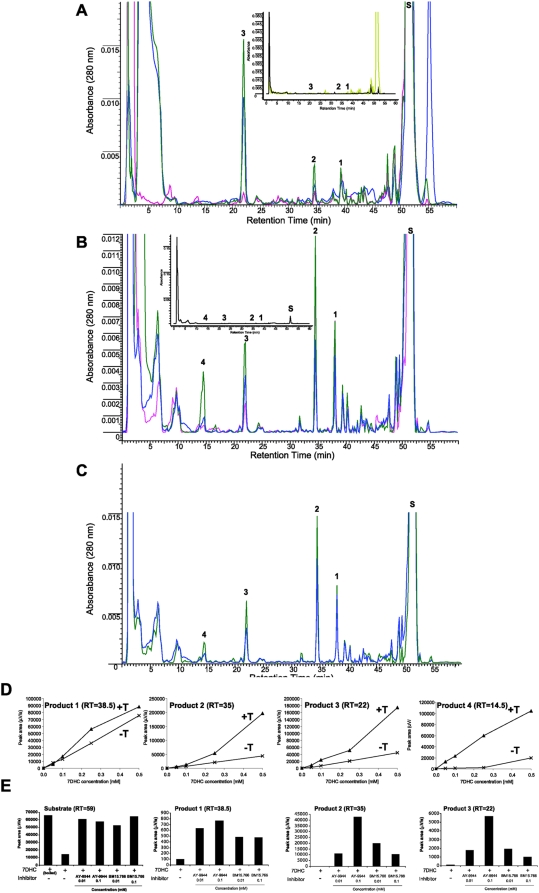
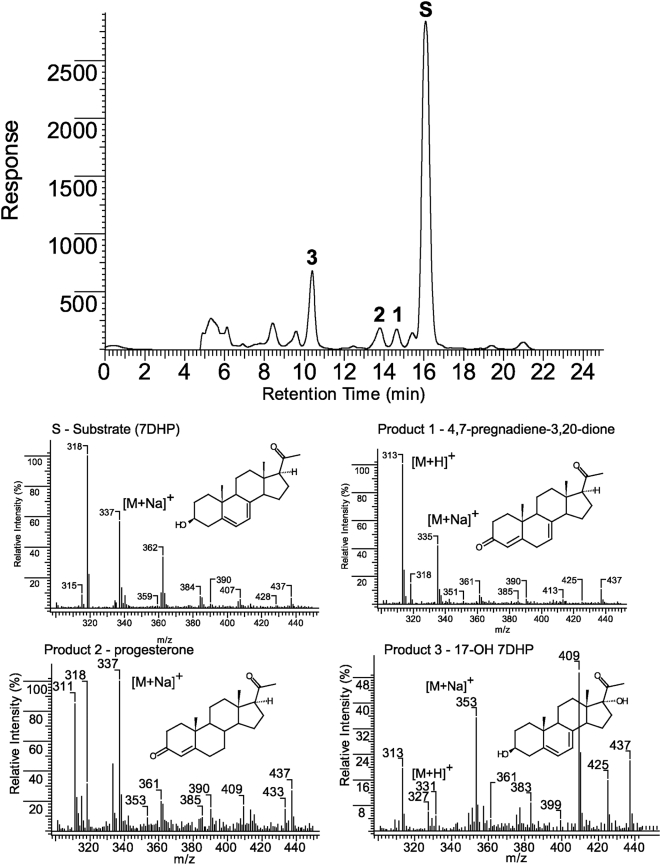
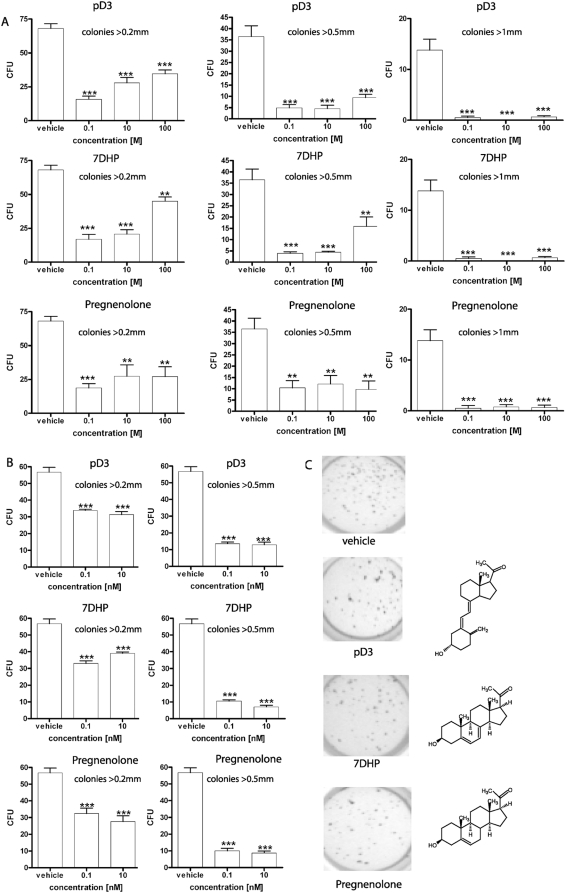
Since P450scc transforms 7-dehydrocholesterol (7DHC) to 7-dehydropregnenolone (7DHP) in vitro, we investigated sequential 7DHC metabolism by adrenal glands ex vivo. There was a rapid, time- and dose-dependent metabolism of 7DHC by adrenals from rats, pigs, rabbits and dogs with production of more polar 5,7-dienes as detected by RP-HPLC. Based on retention time (RT), UV spectra and mass spectrometry, we identified the major products common to all tested species as 7DHP, 22-hydroxy-7DHC and 20,22-dihydroxy-7DHC. The involvement of P450scc in adrenal metabolic transformation was confirmed by the inhibition of this process by DL-aminoglutethimide. The metabolism of 7DHC with subsequent production of 7DHP was stimulated by forscolin indicating involvement of cAMP dependent pathways. Additional minor products of 7DHC metabolism that were more polar than 7DHP were identified as 17-hydroxy-7DHP (in pig adrenals but not those of rats) and as pregna-4,7-diene-3,20-dione (7-dehydroprogesterone). Both products represented the major identifiable products of 7DHP metabolism in adrenal glands. Studies with purified enzymes show that StAR protein likely transports 7DHC to the inner mitochondrial membrane, that 7DHC can compete effectively with cholesterol for the substrate binding site on P450scc and that the catalytic efficiency of 3betaHSD for 7DHP (V(m)/K(m)) is 40% of that for pregnenolone. Skin mitochondria are capable of transforming 7DHC to 7DHP and the 7DHP is metabolized further by skin extracts. Finally, 7DHP, its photoderivative 20-oxopregnacalciferol, and pregnenolone exhibited biological activity in skin cells including inhibition of proliferation of epidermal keratinocytes and melanocytes, and melanoma cells. These findings define a novel steroidogenic pathway: 7DHC-->22(OH)7DHC-->20,22(OH)(2)7DHC-->7DHP, with potential further metabolism of 7DHP mediated by 3betaHSD or CYP17, depending on mammalian species. The 5-7 dienal intermediates of the pathway can be a source of biologically active vitamin D3 derivatives after delivery to or production in the skin, an organ intermittently exposed to solar radiation.
Conflict of interest statement
Figures








References
-
- Payne AH, Hales DB. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr Rev. 2004;25:947–70. - PubMed
-
- Tuckey RC. Progesterone synthesis by the human placenta. Placenta. 2005;26:273–81. - PubMed
-
- Burstein S, Gut M. Intermediates in the conversion of cholesterol to pregnenolone: kinetics and mechanism. Steroids. 1976;28:115–31. - PubMed
-
- Hume R, Kelly RW, Taylor PL, Boyd GS. The catalytic cycle of cytochrome P-450scc and intermediates in the conversion of cholesterol to pregnenolone. Eur J Biochem. 1984;140:583–91. - PubMed
-
- Lambeth JD, Seybert DW, Lancaster JR, Jr, Salerno JC, Kamin H. Steroidogenic electron transport in adrenal cortex mitochondria. Mol Cell Biochem. 1982;45:13–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
