

Effect of bone morphogenetic protein-6 on macrophages
- PMID: 19191909
- PMCID: PMC2753950
- DOI: 10.1111/j.1365-2567.2008.02998.x
Effect of bone morphogenetic protein-6 on macrophages
Abstract
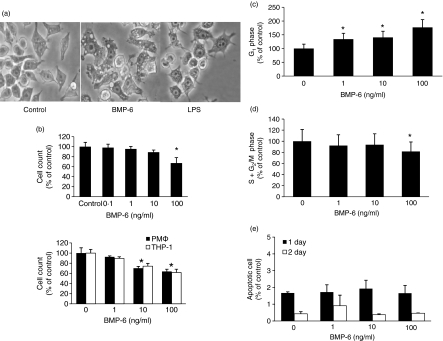
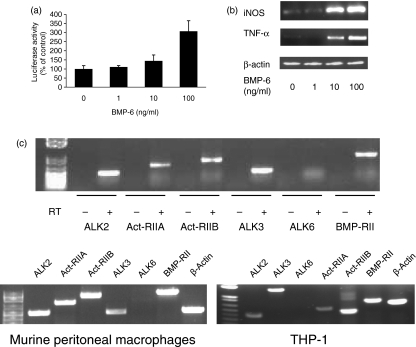
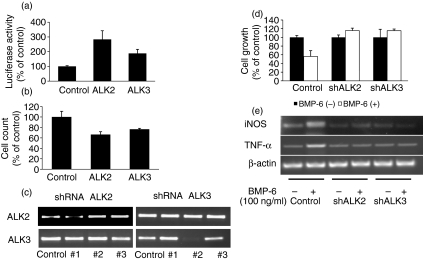
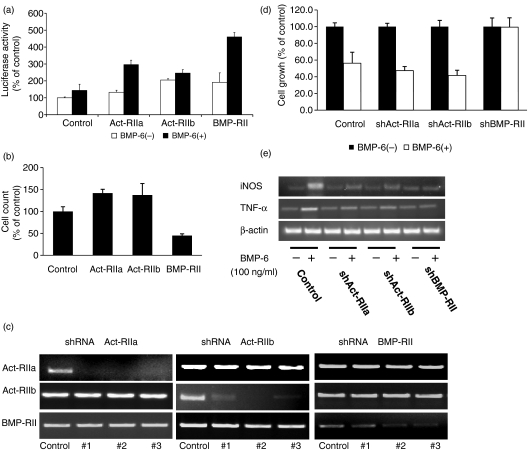
Bone morphogenetic proteins (BMPs) are members of the transforming growth factor (TGF)-beta superfamily which regulates bone formation, haematopoiesis and development. While TGF-beta is known to be a negative regulator of the immune system, the effect of BMPs on the immune system is largely unknown. Herein, the effect of BMP-6 on the innate immune system was investigated using the murine macrophage cell line RAW 264.7. BMP-6 altered cellular morphology, inhibited cellular proliferation, increased the fraction of subG(1) phase cells, and decreased the fraction of cells in the S and G(2)M phases, without changing the percentage of apoptotic cells. In addition, BMP-6 induced expression of pro-inflammatory inducible nitric oxide synthase (iNOS) and the cytokine tumour necrosis factor (TNF)-alpha. Reverse transcription-polymerase chain reaction (RT-PCR) analysis demonstrated the expression of all three known type II BMP receptors [BMP-RII, activin (Act)-RIIA and Act-RIIB] and two of the three known type I receptors [activin receptor-like kinase 2 (ALK2) and ALK3]. Over-expression as well as knock-down studies using short hairpin RNA (shRNA) demonstrated that BMP-RII, ALK2 and ALK3 are the functional BMP-6 receptors in macrophages. Finally, the effect of BMP-6 was confirmed in murine peritoneal macrophages and the THP-1 human monocyte cell line. Taken together, these results demonstrate that BMP-6 regulates the proliferation and gene expression profile of macrophages.
Figures
References
-
- Wozney JM, Rosen V, Celeste AJ, Mitsock LM, Whitters MJ, Kriz RW, Hewick RM, Wang EA. Novel regulators of bone formation: molecular clones and activities. Science. 1988;242:1528–34. - PubMed
-
- Hogan BL. Bone morphogenetic proteins: multifunctional regulators of vertebrate development. Genes Dev. 1996;10:1580–94. - PubMed
-
- Zhao GQ. Consequences of knocking out BMP signaling in the mouse. Genesis. 2003;35:43–56. - PubMed
-
- Sivertsen EA, Huse K, Hystad ME, Kersten C, Smeland EB, Myklebust JH. Inhibitory effects and target genes of bone morphogenetic protein 6 in Jurkat TAg cells. Eur J Immunol. 2007;37:2937–48. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
