

Basal body assembly in ciliates: the power of numbers
- PMID: 19192246
- PMCID: PMC2743681
- DOI: 10.1111/j.1600-0854.2009.00885.x
Basal body assembly in ciliates: the power of numbers
Abstract
Centrioles perform the dual functions of organizing both centrosomes and cilia. The biogenesis of nascent centrioles is an essential cellular event that is tightly coupled to the cell cycle so that each cell contains only two or four centrioles at any given point in the cell cycle. The assembly of centrioles and their analogs, basal bodies, is well characterized at the ultrastructural level whereby structural modules are built into a functional organelle. Genetic studies in model organisms combined with proteomic, bioinformatic and identifying ciliary disease gene orthologs have revealed a wealth of molecules requiring further analysis to determine their roles in centriole duplication, assembly and function. Nonetheless, at this stage, our understanding of how molecular components interact to build new centrioles and basal bodies is limited. The ciliates, Tetrahymena and Paramecium, historically have been the subject of cytological and genetic study of basal bodies. Recent advances in the ciliate genetic and molecular toolkit have placed these model organisms in a favorable position to study the molecular mechanisms of centriole and basal body assembly.
Figures
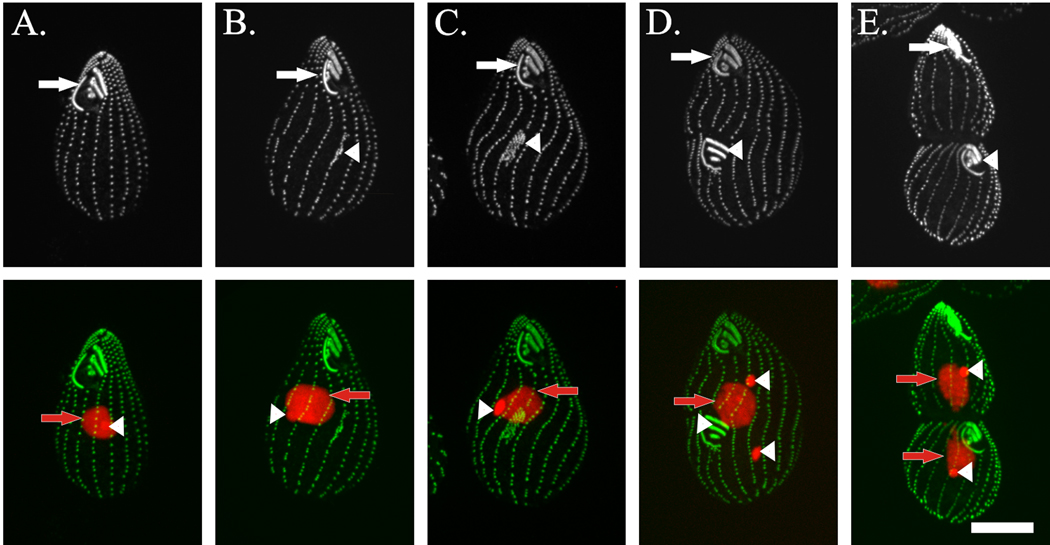
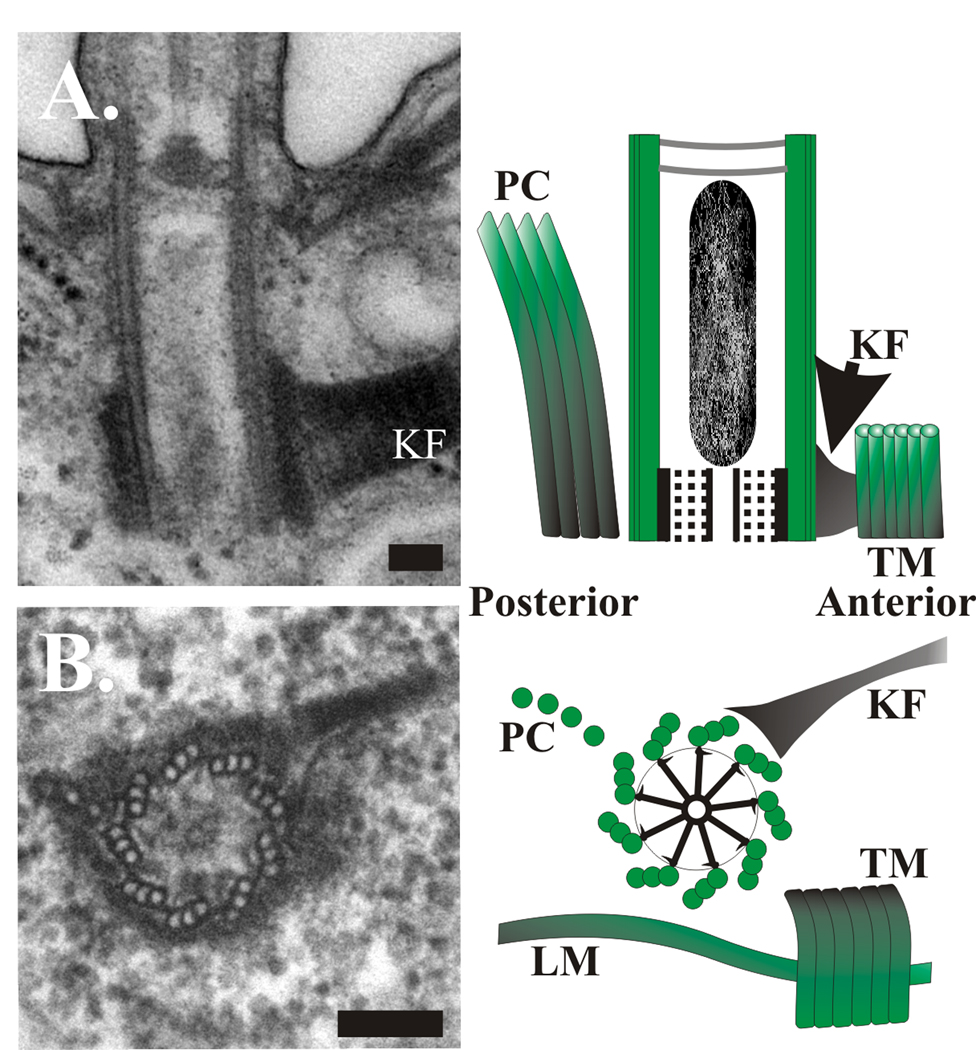
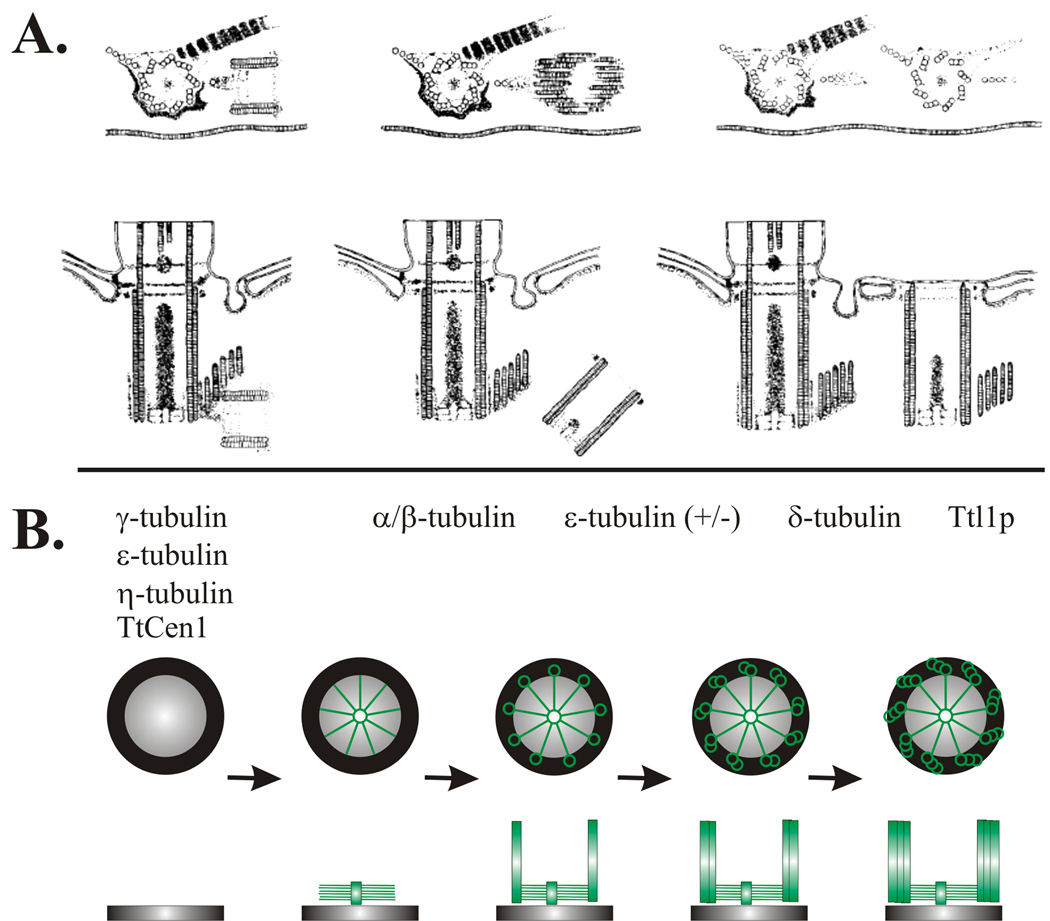
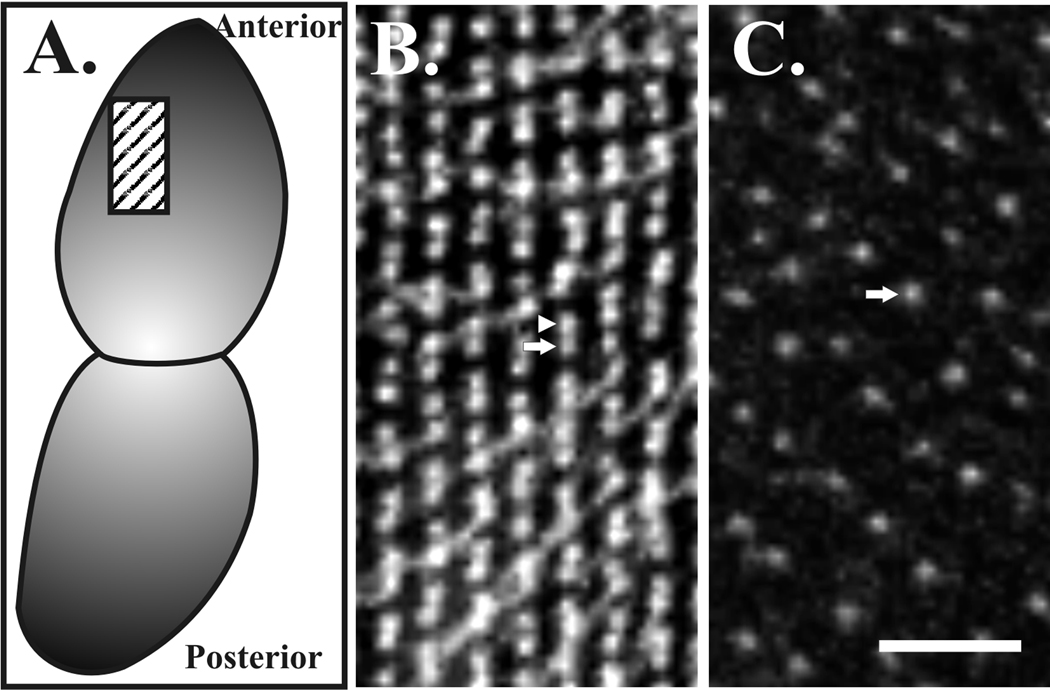
References
-
- Boveri T. Germany: Jena; 1914. Zur frage der entstehung maligner tumoren.
-
- Wilson E. 3rd ed. New York: Macmillan; 1925. The Cell in Development and Heredity.
-
- Bornens M, Azimzadeh J. Origin and evolution of the centrosome. Advances in experimental medicine and biology. 2007;607:119–129. - PubMed
-
- Basto R, Lau J, Vinogradova T, Gardiol A, Woods CG, Khodjakov A, Raff JW. Flies without centrioles. Cell. 2006;125(7):1375–1386. - PubMed
-
- Marshall WF, Nonaka S. Cilia: tuning in to the cell's antenna. Curr Biol. 2006;16(15):R604–R614. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
