

Hyaluronan orchestrates transforming growth factor-beta1-dependent maintenance of myofibroblast phenotype
- PMID: 19193641
- PMCID: PMC2666557
- DOI: 10.1074/jbc.M806989200
Hyaluronan orchestrates transforming growth factor-beta1-dependent maintenance of myofibroblast phenotype
Abstract
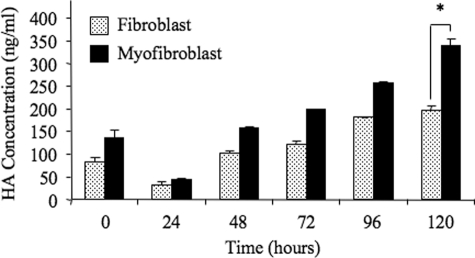
The differentiation of resident fibroblasts to myofibroblasts is central to wound healing. In the context of organ fibrosis, however, persistence of these myofibroblasts is associated with progressive disease. This study examines mechanisms controlling the maintenance of the myofibroblast phenotype. Myofibroblasts were induced by adding transforming growth factor-beta1 (TGF-beta1) (10 ng/ml) to fibroblasts for 72 h. The phenotype was maintained for up to 120 h following removal of TGF-beta1. Western blot for pSmad2 and -3 demonstrated persistent phosphorylation despite removal of exogenous TGF-beta1. This persistence was because of autocrine synthesis of TGF-beta1, which was inhibited by both anti-TGF-beta1 antibody and the ALK5 inhibitor SB431542. Persistence of phenotype was also associated with increased hyaluronan (HA) generation, synthesis of the hyaladherin TSG6, and HA pericellular coat formation. These were all inhibited by TGF-beta receptor blockade. To further investigate the importance of HA synthesis, 4-methylumbelliferone was used to deplete the cytoplasmic pool of UDP-glucuronic acid, essential for HA chain elongation. This prevented formation of the pericellular HA matrix and decreased expression of alpha-SMA. 4-Methylumbelliferone had no effect, however, on Smad2 and -3 phosphorylation. Similarly inhibition of HAS2 by short interfering RNA prevented phenotypic activation without altering TGF-beta1-dependent Smad phosphorylation, thus suggesting that HA-dependent regulation of cell phenotype was independent of Smad activation. These data suggest that myofibroblasts in areas of fibrosis maintain their own phenotype through autocrine TGF-beta1 action and that extracellular HA matrices are an essential mediator of this. We propose a model in which the formation of the pericellular HA matrix regulates the outcome of Smad-dependent autocrine TGF-beta1-activated signaling, and therefore persistence of the myofibroblast phenotype.
Figures








References
-
- Sappino, A. P., Schürch, W., and Gabbiani, G. (1990) Lab. Investig. 63 144–161 - PubMed
-
- Gabbiani, G. (2003) J. Pathol. 200 500–503 - PubMed
-
- Desmouliere, A., and Gabbiani, G. (1996) in The Molecular and Cellular Biology of Wound Repair (Clark, R. A. F., ed) 2nd Ed., pp. 391–423, Plenum Publishing Corp., New York
-
- Essawy, M., Soylemezoglu, O., Muchaneta-Kubara, E. C., Shortland, J., Brown, C. B., and El Nahas, A. M. (1997) Nephrol. Dial. Transplant. 12 43–50 - PubMed
-
- Goumenos, D. S., Brown, C. B., Shortland, J., and El Nahas, A. M. (1994) Nephrol. Dial. Transplant. 9 1418–1425 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
