

At least ten genes define the imprinted Dlk1-Dio3 cluster on mouse chromosome 12qF1
- PMID: 19194500
- PMCID: PMC2632752
- DOI: 10.1371/journal.pone.0004352
At least ten genes define the imprinted Dlk1-Dio3 cluster on mouse chromosome 12qF1
Abstract
Background: Genomic imprinting is an exception to Mendelian genetics in that imprinted genes are expressed monoallelically, dependent on parental origin. In mammals, imprinted genes are critical in numerous developmental and physiological processes. Aberrant imprinted gene expression is implicated in several diseases including Prader-Willi/Angelman syndromes and cancer.
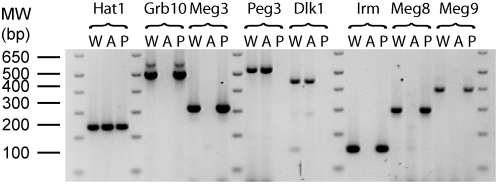
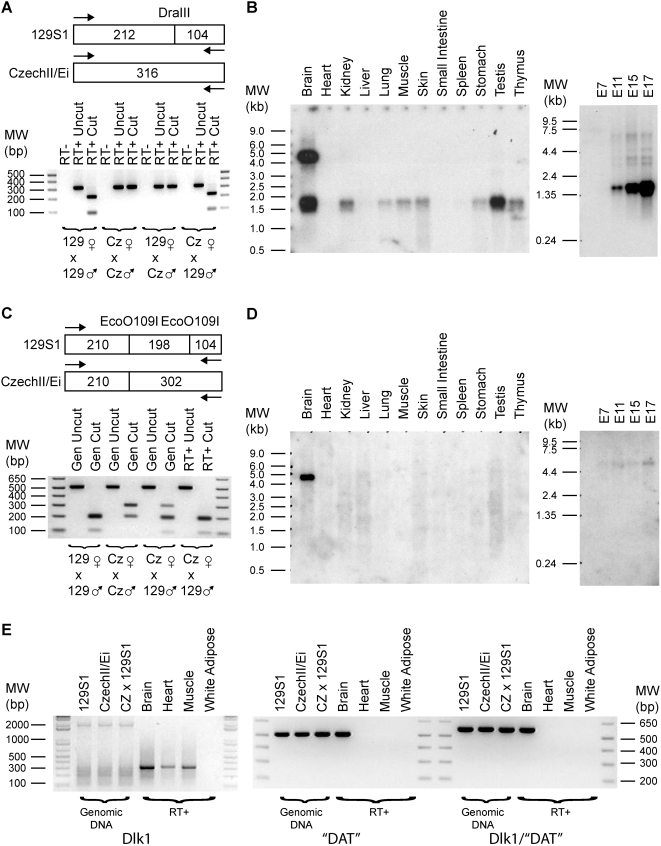
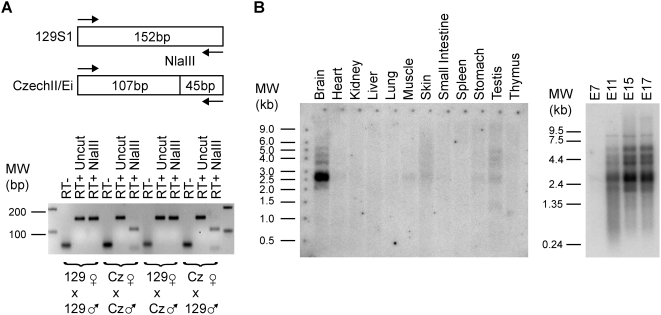
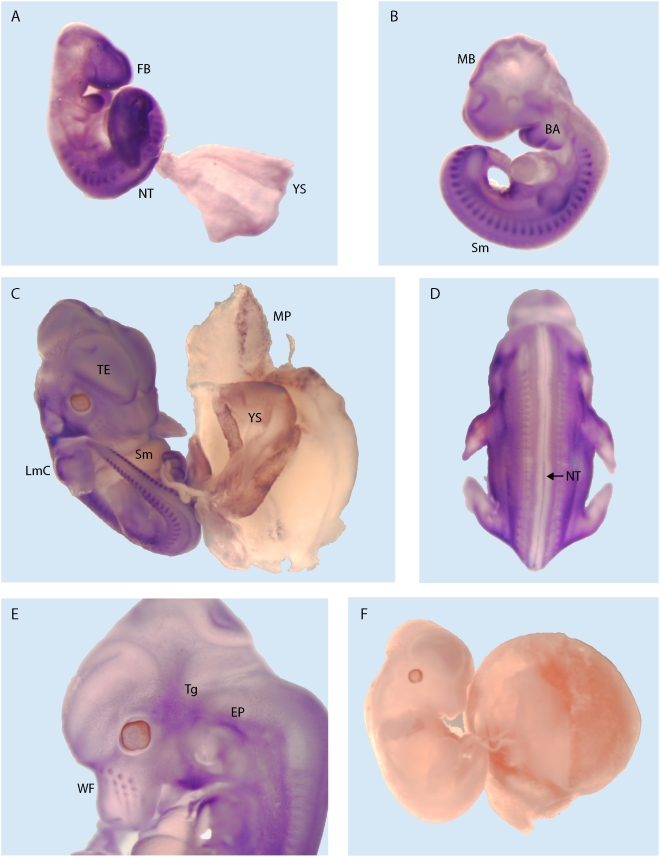
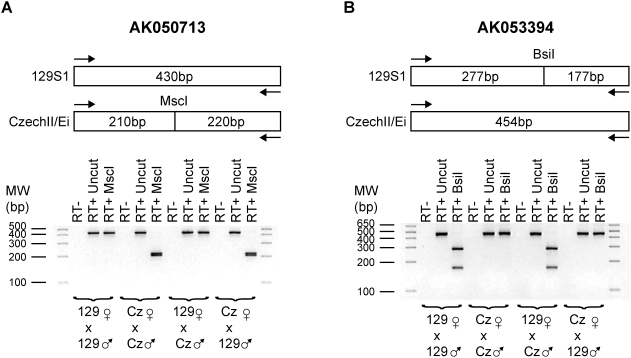
Methodology/principal findings: To identify novel imprinted genes, transcription profiling was performed on two uniparentally derived cell lines, androgenetic and parthenogenetic primary mouse embryonic fibroblasts. A maternally expressed transcript termed Imprinted RNA near Meg3/Gtl2 (Irm) was identified and its expression studied by Northern blotting and whole mounts in situ hybridization. The imprinted region that contains Irm has a parent of origin effect in three mammalian species, including the sheep callipyge locus. In mice and humans, both maternal and paternal uniparental disomies (UPD) cause embryonic growth and musculoskeletal abnormalities, indicating that both alleles likely express essential genes. To catalog all imprinted genes in this chromosomal region, twenty-five mouse mRNAs in a 1.96Mb span were investigated for allele specific expression.
Conclusions/significance: Ten imprinted genes were elucidated. The imprinting of three paternally expressed protein coding genes (Dlk1, Peg11, and Dio3) was confirmed. Seven noncoding RNAs (Meg3/Gtl2, Anti-Peg11, Meg8, Irm/"Rian", AK050713, AK053394, and Meg9/Mirg) are characterized by exclusive maternal expression. Intriguingly, the majority of these noncoding RNA genes contain microRNAs and/or snoRNAs within their introns, as do their human orthologs. Of the 52 identified microRNAs that map to this region, six are predicted to regulate negatively Dlk1, suggesting an additional mechanism for interactions between allelic gene products. Since several previous studies relied heavily on in silico analysis and RT-PCR, our findings from Northerns and cDNA cloning clarify the genomic organization of this region. Our results expand the number of maternally expressed noncoding RNAs whose loss may be responsible for the phenotypes associated with mouse pUPD12 and human pUPD14 syndromes.
Conflict of interest statement
Figures






Similar articles
-
[Review on the genomic imprinting at the mammalian DLK1-DIO3 cluster.].Yi Chuan. 2010 Aug;32(8):769-78. doi: 10.3724/sp.j.1005.2010.00769. Yi Chuan. 2010. PMID: 20709673 Review. Chinese.
-
Mouse Peg9/Dlk1 and human PEG9/DLK1 are paternally expressed imprinted genes closely located to the maternally expressed imprinted genes: mouse Meg3/Gtl2 and human MEG3.Genes Cells. 2000 Dec;5(12):1029-37. doi: 10.1046/j.1365-2443.2000.00390.x. Genes Cells. 2000. PMID: 11168589
-
Dlk1-Dio3 locus-derived lncRNAs perpetuate postmitotic motor neuron cell fate and subtype identity.Elife. 2018 Oct 12;7:e38080. doi: 10.7554/eLife.38080. Elife. 2018. PMID: 30311912 Free PMC article.
-
Differential regulation of imprinting in the murine embryo and placenta by the Dlk1-Dio3 imprinting control region.Development. 2007 Jan;134(2):417-26. doi: 10.1242/dev.02726. Epub 2006 Dec 13. Development. 2007. PMID: 17166925
-
The microRNAs within the DLK1-DIO3 genomic region: involvement in disease pathogenesis.Cell Mol Life Sci. 2013 Mar;70(5):795-814. doi: 10.1007/s00018-012-1080-8. Epub 2012 Jul 24. Cell Mol Life Sci. 2013. PMID: 22825660 Free PMC article. Review.
Cited by
-
Upregulation of the microRNA cluster at the Dlk1-Dio3 locus in lung adenocarcinoma.Oncogene. 2015 Jan 2;34(1):94-103. doi: 10.1038/onc.2013.523. Epub 2013 Dec 9. Oncogene. 2015. PMID: 24317514 Free PMC article.
-
Maternal inheritance of an inactive type III deiodinase gene allele affects mouse pancreatic β-cells and disrupts glucose homeostasis.Endocrinology. 2014 Aug;155(8):3160-71. doi: 10.1210/en.2013-1208. Epub 2014 Jun 2. Endocrinology. 2014. PMID: 24885572 Free PMC article.
-
Tissue-specific alternative polyadenylation at the imprinted gene Mest regulates allelic usage at Copg2.Nucleic Acids Res. 2012 Feb;40(4):1523-35. doi: 10.1093/nar/gkr871. Epub 2011 Nov 3. Nucleic Acids Res. 2012. PMID: 22053079 Free PMC article.
-
Parent-of-origin effects implicate epigenetic regulation of experimental autoimmune encephalomyelitis and identify imprinted Dlk1 as a novel risk gene.PLoS Genet. 2014 Mar 27;10(3):e1004265. doi: 10.1371/journal.pgen.1004265. eCollection 2014 Mar. PLoS Genet. 2014. PMID: 24676147 Free PMC article.
-
Regulatory mechanisms of long noncoding RNAs in vertebrate central nervous system development and function.Neuroscience. 2013 Apr 3;235:200-14. doi: 10.1016/j.neuroscience.2013.01.022. Epub 2013 Jan 18. Neuroscience. 2013. PMID: 23337534 Free PMC article. Review.
References
-
- McGrath J, Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell. 1984;37:179–183. - PubMed
-
- Barton SC, Surani MA, Norris ML. Role of paternal and maternal genomes in mouse development. Nature. 1984;311:374–376. - PubMed
-
- Esteller M, Herman JG. Cancer as an epigenetic disease: DNA methylation and chromatin alterations in human tumours. J Pathol. 2002;196:1–7. - PubMed
-
- Ogawa O, Eccles MR, Szeto J, McNoe LA, Yun K, et al. Relaxation of insulin-like growth factor II gene imprinting implicated in Wilms' tumour. Nature. 1993;362:749–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
