

Linker histone H1 is essential for Drosophila development, the establishment of pericentric heterochromatin, and a normal polytene chromosome structure
- PMID: 19196654
- PMCID: PMC2648648
- DOI: 10.1101/gad.1749309
Linker histone H1 is essential for Drosophila development, the establishment of pericentric heterochromatin, and a normal polytene chromosome structure
Abstract
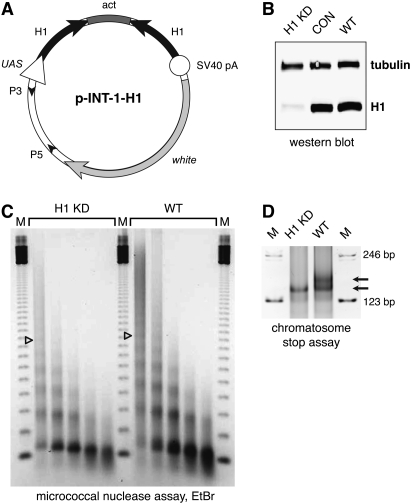
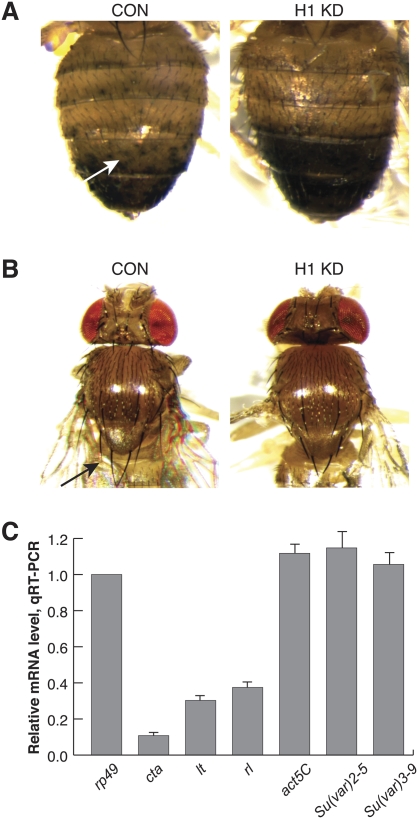
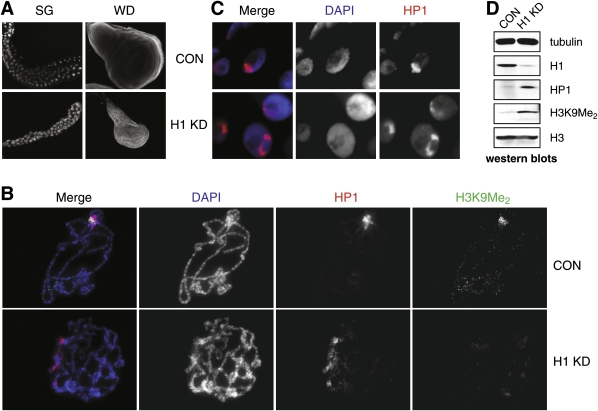
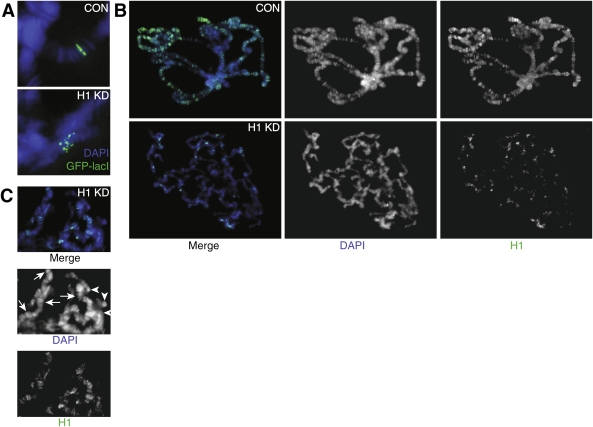
We generated mutant alleles of Drosophila melanogaster in which expression of the linker histone H1 can be down-regulated over a wide range by RNAi. When the H1 protein level is reduced to approximately 20% of the level in wild-type larvae, lethality occurs in the late larval - pupal stages of development. Here we show that H1 has an important function in gene regulation within or near heterochromatin. It is a strong dominant suppressor of position effect variegation (PEV). Similar to other suppressors of PEV, H1 is simultaneously involved in both the repression of euchromatic genes brought to the vicinity of pericentric heterochromatin and the activation of heterochromatic genes that depend on their pericentric localization for maximal transcriptional activity. Studies of H1-depleted salivary gland polytene chromosomes show that H1 participates in several fundamental aspects of chromosome structure and function. First, H1 is required for heterochromatin structural integrity and the deposition or maintenance of major pericentric heterochromatin-associated histone marks, including H3K9Me(2) and H4K20Me(2). Second, H1 also plays an unexpected role in the alignment of endoreplicated sister chromatids. Finally, H1 is essential for organization of pericentric regions of all polytene chromosomes into a single chromocenter. Thus, linker histone H1 is essential in Drosophila and plays a fundamental role in the architecture and activity of chromosomes in vivo.
Figures
References
-
- Ananiev E.V., Barsky V.E. Elementary structures in polytene chromosomes of Drosophila melanogaster. Chromosoma. 1985;93:104–112. - PubMed
-
- Brand A.H., Manoukian A.S., Perrimon N. Ectopic expression in Drosophila. Methods Cell Biol. 1994;44:635–654. - PubMed
-
- Cartwright I.L., Cryderman D.E., Gilmour D.S., Pile L.A., Wallrath L.L., Weber J.A., Elgin S.C. Analysis of Drosophila chromatin structure in vivo. Methods Enzymol. 1999;304:462–496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases