

Calcium-activated nonspecific cation current and synaptic depression promote network-dependent burst oscillations
- PMID: 19196976
- PMCID: PMC2636730
- DOI: 10.1073/pnas.0808776106
Calcium-activated nonspecific cation current and synaptic depression promote network-dependent burst oscillations
Abstract
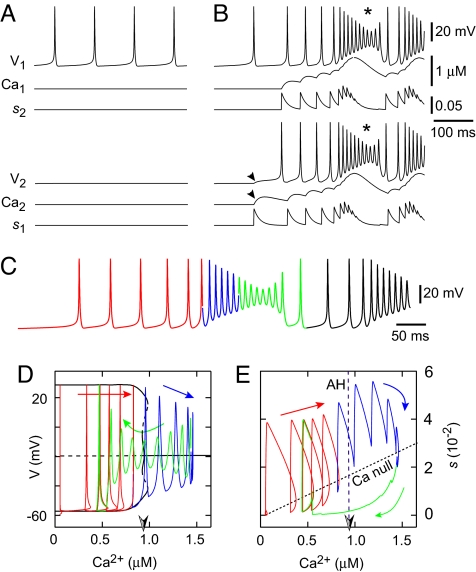
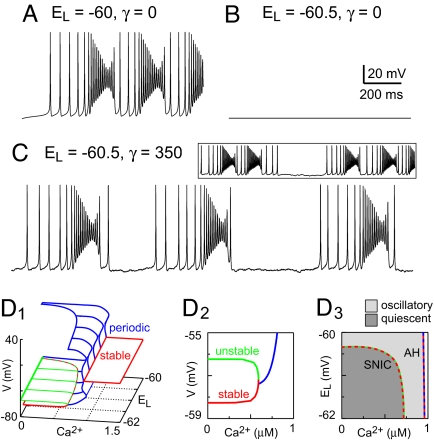
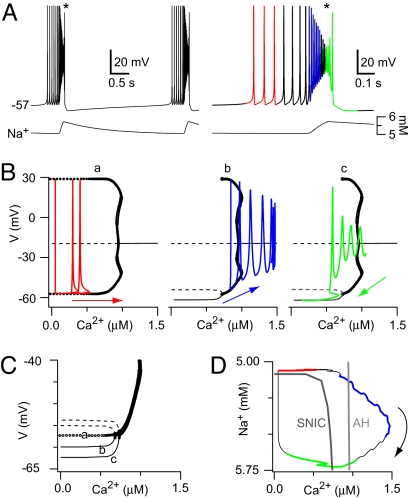
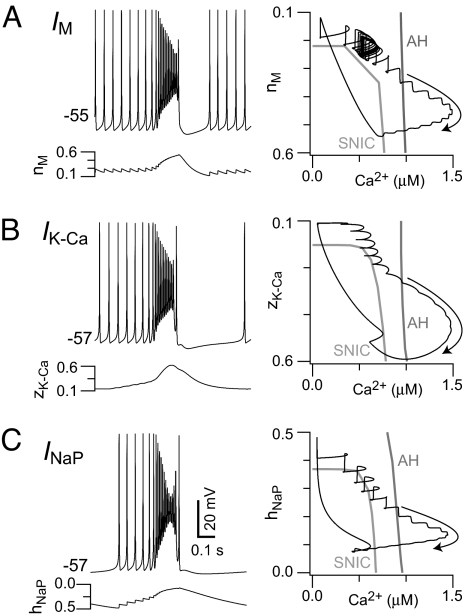
Central pattern generators (CPGs) produce neural-motor rhythms that often depend on specialized cellular or synaptic properties such as pacemaker neurons or alternating phases of synaptic inhibition. Motivated by experimental evidence suggesting that activity in the mammalian respiratory CPG, the preBötzinger complex, does not require either of these components, we present and analyze a mathematical model demonstrating an unconventional mechanism of rhythm generation in which glutamatergic synapses and the short-term depression of excitatory transmission play key rhythmogenic roles. Recurrent synaptic excitation triggers postsynaptic Ca(2+)-activated nonspecific cation current (I(CAN)) to initiate a network-wide burst. Robust depolarization due to I(CAN) also causes voltage-dependent spike inactivation, which diminishes recurrent excitation and thus attenuates postsynaptic Ca(2+) accumulation. Consequently, activity-dependent outward currents-produced by Na/K ATPase pumps or other ionic mechanisms-can terminate the burst and cause a transient quiescent state in the network. The recovery of sporadic spiking activity rekindles excitatory interactions and initiates a new cycle. Because synaptic inputs gate postsynaptic burst-generating conductances, this rhythm-generating mechanism represents a new paradigm that can be dubbed a 'group pacemaker' in which the basic rhythmogenic unit encompasses a fully interdependent ensemble of synaptic and intrinsic components. This conceptual framework should be considered as an alternative to traditional models when analyzing CPGs for which mechanistic details have not yet been elucidated.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Local mechanisms of phase-dependent postsynaptic modifications of NMDA-induced oscillations in the abducens motoneurons: a simulation study.J Neurophysiol. 1996 Aug;76(2):1015-24. doi: 10.1152/jn.1996.76.2.1015. J Neurophysiol. 1996. PMID: 8871216
-
Computer simulations of NMDA and non-NMDA receptor-mediated synaptic drive: sensory and supraspinal modulation of neurons and small networks.J Neurophysiol. 1993 Aug;70(2):695-709. doi: 10.1152/jn.1993.70.2.695. J Neurophysiol. 1993. PMID: 8105036
-
Synaptically activated burst-generating conductances may underlie a group-pacemaker mechanism for respiratory rhythm generation in mammals.Prog Brain Res. 2010;187:111-36. doi: 10.1016/B978-0-444-53613-6.00008-3. Prog Brain Res. 2010. PMID: 21111204 Free PMC article.
-
The impact of single cell voltage clamp on the understanding of the cardiac ventricular action potential.Cardioscience. 1992 Sep;3(3):131-44. Cardioscience. 1992. PMID: 1384746 Review.
-
Neuronal excitability: voltage-dependent currents and synaptic transmission.J Clin Neurophysiol. 1992 Apr;9(2):195-211. J Clin Neurophysiol. 1992. PMID: 1375602 Review.
Cited by
-
Dual oscillator model of the respiratory neuronal network generating quantal slowing of respiratory rhythm.J Comput Neurosci. 2011 Apr;30(2):225-40. doi: 10.1007/s10827-010-0249-0. Epub 2010 Jun 11. J Comput Neurosci. 2011. PMID: 20544264 Free PMC article.
-
Genetic identification of an embryonic parafacial oscillator coupling to the preBötzinger complex.Nat Neurosci. 2009 Aug;12(8):1028-35. doi: 10.1038/nn.2354. Epub 2009 Jul 5. Nat Neurosci. 2009. PMID: 19578380
-
Synaptic Depression Influences Inspiratory-Expiratory Phase Transition in Dbx1 Interneurons of the preBötzinger Complex in Neonatal Mice.J Neurosci. 2015 Aug 19;35(33):11606-11. doi: 10.1523/JNEUROSCI.0351-15.2015. J Neurosci. 2015. PMID: 26290237 Free PMC article.
-
Evolution of vocal patterns: tuning hindbrain circuits during species divergence.J Exp Biol. 2017 Mar 1;220(Pt 5):856-867. doi: 10.1242/jeb.146845. Epub 2016 Dec 23. J Exp Biol. 2017. PMID: 28011819 Free PMC article.
-
Functional anatomical evidence for respiratory rhythmogenic function of endogenous bursters in rat medulla.J Neurosci. 2010 Jun 23;30(25):8383-92. doi: 10.1523/JNEUROSCI.5510-09.2010. J Neurosci. 2010. PMID: 20573885 Free PMC article.
References
-
- Gosgnach S, et al. V1 spinal neurons regulate the speed of vertebrate locomotor outputs. Nature. 2006;440:215–219. - PubMed
-
- Marder E. Moving rhythms. Nature. 2001;410:755. - PubMed
-
- Grillner S. Biological pattern generation: The cellular and computational logic of networks in motion. Neuron. 2006;52:751–766. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
