

Wnt signaling, lgr5, and stem cells in the intestine and skin
- PMID: 19197002
- PMCID: PMC2665733
- DOI: 10.2353/ajpath.2009.080758
Wnt signaling, lgr5, and stem cells in the intestine and skin
Abstract
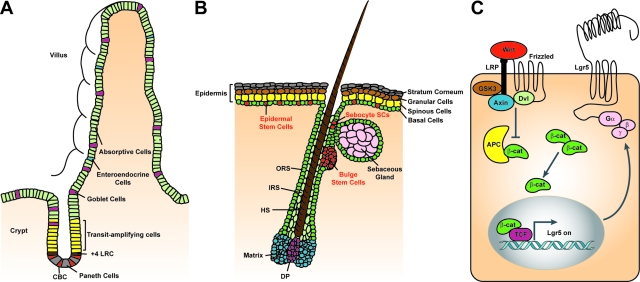
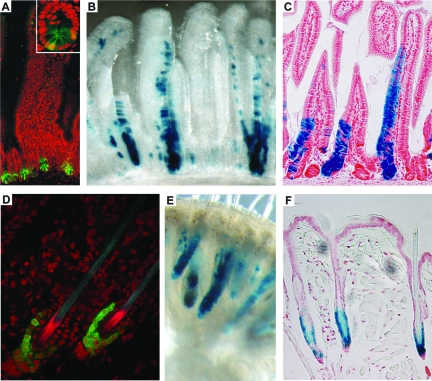
Stem cells hold great promise for regenerative medicine, but have remained elusive in many tissues because of a lack of adequate definitive markers. Progress in mouse genetics has provided the tools for characterization and validation of stem cell markers by functional and/or lineage tracing assays. The Wnt target gene Lgr5 has been recently identified as a novel stem cell marker of the intestinal epithelium and the hair follicle. In the intestine, Lgr5 is exclusively expressed in cycling crypt base columnar cells. Genetic lineage-tracing experiments revealed that crypt base columnar cells are capable of self-renewal and multipotency, thus representing genuine intestinal stem cells. In the stem cell niche of the murine hair follicle, Lgr5 is expressed in actively cycling cells. Transplantation and lineage tracing experiments have demonstrated that these Lgr5(+ve) cells maintain all cell lineages of the hair follicle throughout long periods of time and can build entire new hair follicles. Expression of Lgr5 in multiple other organs indicates that it may represent a global marker of adult stem cells. This review attempts to provide a comprehensive overview of the stem cell compartments in the intestine and skin with a focus on the cycling, yet long-lived and multipotent, Lgr5(+ve) stem cell populations.
Figures
References
-
- Kiel MJ, Yilmaz OH, Iwashita T, Yilmaz OH, Terhorst C, Morrison SJ. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005;121:1109–1121. - PubMed
-
- Shackleton M, Vaillant F, Simpson KJ, Stingl J, Smyth GK, Asselin-Labat ML, Wu L, Lindeman GJ, Visvader JE. Generation of a functional mammary gland from a single stem cell. Nature. 2006;439:84–88. - PubMed
-
- Stingl J, Eirew P, Ricketson I, Shackleton M, Vaillant F, Choi D, Li HI, Eaves CJ. Purification and unique properties of mammary epithelial stem cells. Nature. 2006;439:993–997. - PubMed
-
- Cairns J. Mutation selection and the natural history of cancer. Nature. 1975;255:197–200. - PubMed
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den Born M, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
