

Mislocalization of the MRN complex prevents ATR signaling during adenovirus infection
- PMID: 19197236
- PMCID: PMC2666027
- DOI: 10.1038/emboj.2009.15
Mislocalization of the MRN complex prevents ATR signaling during adenovirus infection
Abstract
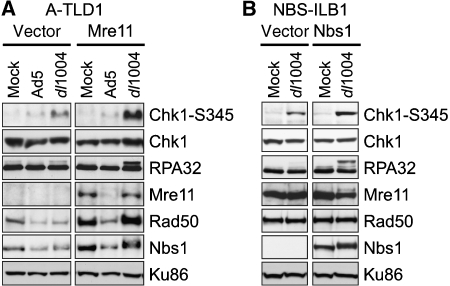
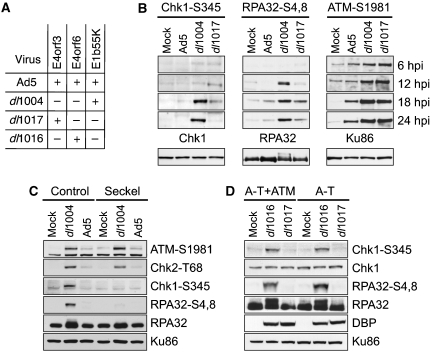
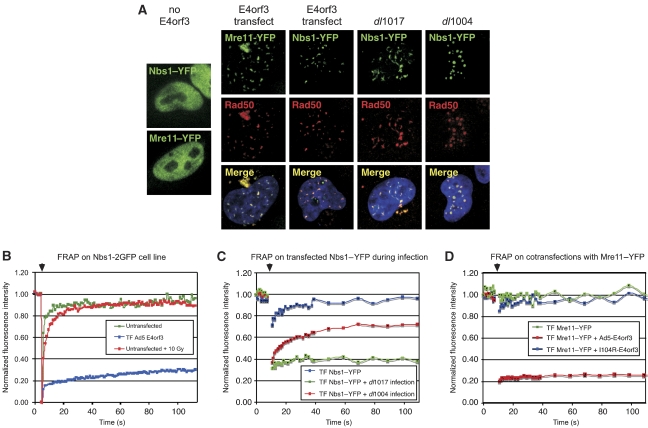
The protein kinases ataxia-telangiectasia mutated (ATM) and ATM-Rad3 related (ATR) are activated in response to DNA damage, genotoxic stress and virus infections. Here we show that during infection with wild-type adenovirus, ATR and its cofactors RPA32, ATRIP and TopBP1 accumulate at viral replication centres, but there is minimal ATR activation. We show that the Mre11/Rad50/Nbs1 (MRN) complex is recruited to viral centres only during infection with adenoviruses lacking the early region E4 and ATR signaling is activated. This suggests a novel requirement for the MRN complex in ATR activation during virus infection, which is independent of Mre11 nuclease activity and recruitment of RPA/ATR/ATRIP/TopBP1. Unlike other damage scenarios, we found that ATM and ATR signaling are not dependent on each other during infection. We identify a region of the viral E4orf3 protein responsible for immobilization of the MRN complex and show that this prevents ATR signaling during adenovirus infection. We propose that immobilization of the MRN damage sensor by E4orf3 protein prevents recognition of viral genomes and blocks detrimental aspects of checkpoint signaling during virus infection.
Figures




References
-
- Arias EE, Walter JC (2007) Strength in numbers: preventing rereplication via multiple mechanisms in eukaryotic cells. Genes Dev 21: 497–518 - PubMed
-
- Bakkenist CJ, Kastan MB (2003) DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421: 499–506 - PubMed
-
- Binz SK, Sheehan AM, Wold MS (2004) Replication protein A phosphorylation and the cellular response to DNA damage. DNA Repair 3: 1015–1024 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
