

High-resolution fMRI of content-sensitive subsequent memory responses in human medial temporal lobe
- PMID: 19199423
- PMCID: PMC2854293
- DOI: 10.1162/jocn.2009.21195
High-resolution fMRI of content-sensitive subsequent memory responses in human medial temporal lobe
Abstract
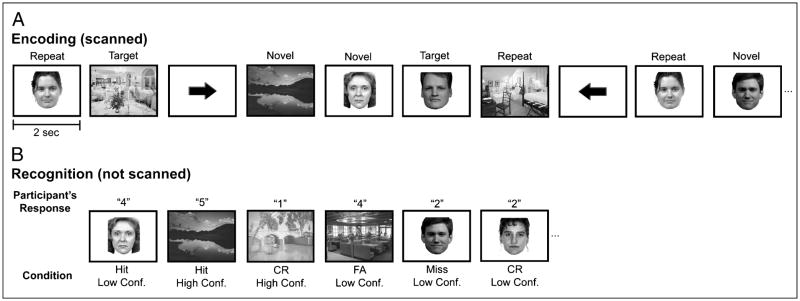
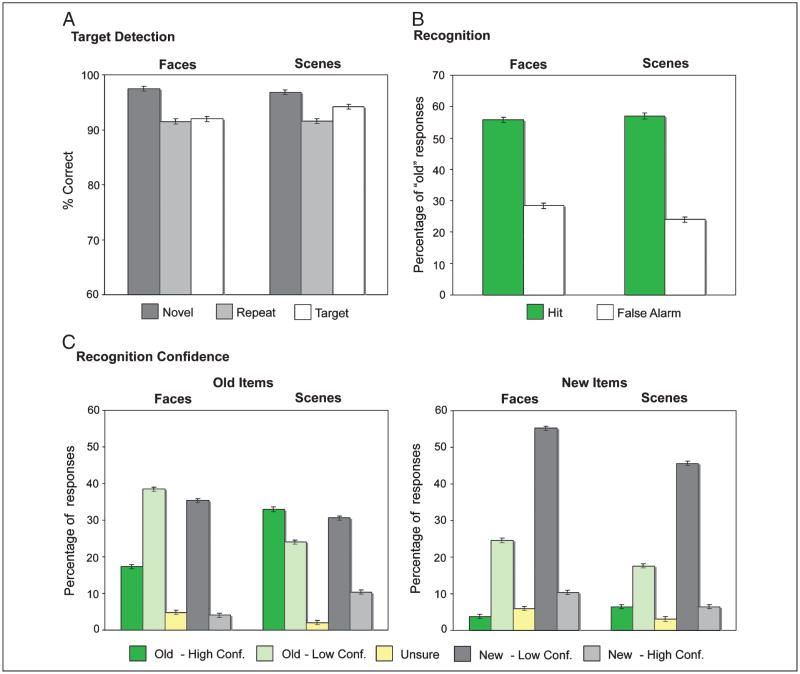
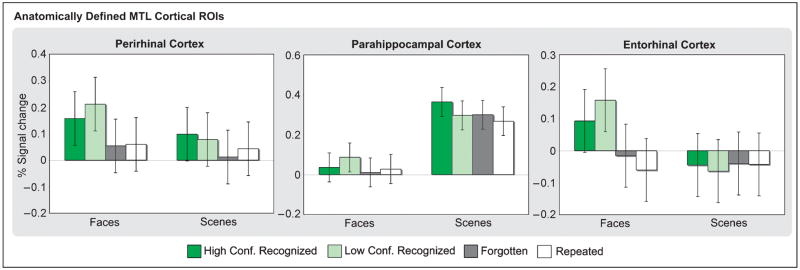
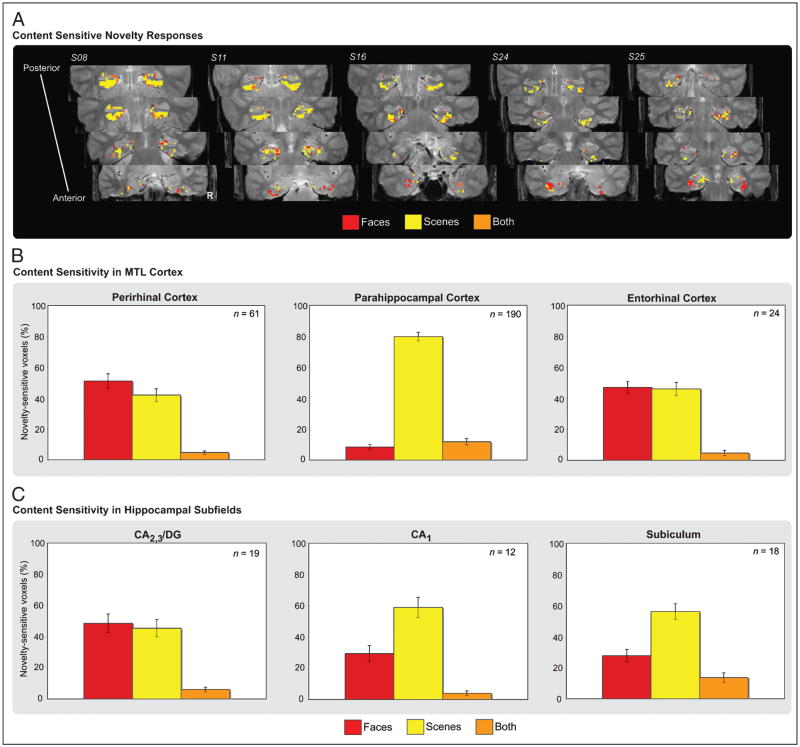
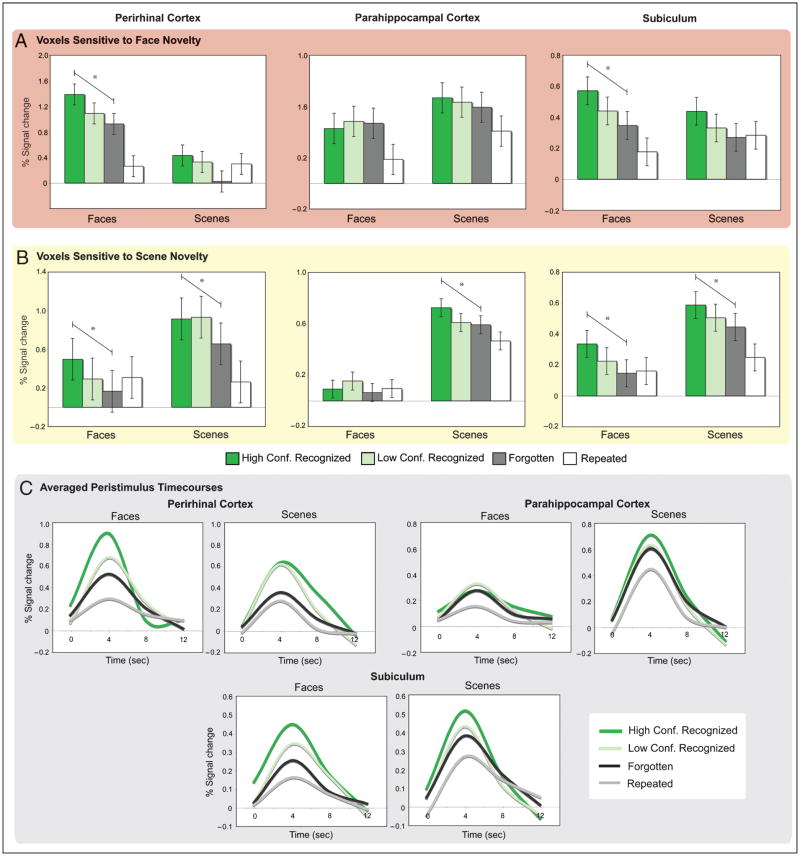
The essential role of the medial temporal lobe (MTL) in long-term memory for individual events is well established, yet important questions remain regarding the mnemonic functions of the component structures that constitute the region. Within the hippocampus, recent functional neuroimaging findings suggest that formation of new memories depends on the dentate gyrus and the CA(3) field, whereas the contribution of the subiculum may be limited to retrieval. During encoding, it has been further hypothesized that structures within MTL cortex contribute to encoding in a content-sensitive manner, whereas hippocampal structures may contribute to encoding in a more domain-general manner. In the current experiment, high-resolution fMRI techniques were utilized to assess novelty and subsequent memory effects in MTL subregions for two classes of stimuli--faces and scenes. During scanning, participants performed an incidental encoding (target detection) task with novel and repeated faces and scenes. Subsequent recognition memory was indexed for the novel stimuli encountered during scanning. Analyses revealed voxels sensitive to both novel faces and novel scenes in all MTL regions. However, similar percentages of voxels were sensitive to novel faces and scenes in perirhinal cortex, entorhinal cortex, and a combined region comprising the dentate gyrus, CA(2), and CA(3), whereas parahippocampal cortex, CA(1), and subiculum demonstrated greater sensitivity to novel scene stimuli. Paralleling these findings, subsequent memory effects in perirhinal cortex were observed for both faces and scenes, with the magnitude of encoding activation being related to later memory strength, as indexed by a graded response tracking recognition confidence, whereas subsequent memory effects were scene-selective in parahippocampal cortex. Within the hippocampus, encoding activation in the subiculum correlated with subsequent memory for both stimulus classes, with the magnitude of encoding activation varying in a graded manner with later memory strength. Collectively, these findings suggest a gradient of content sensitivity from posterior (parahippocampal) to anterior (perirhinal) MTL cortex, with MTL cortical regions differentially contributing to successful encoding based on event content. In contrast to recent suggestions, the present data further indicate that the subiculum may contribute to successful encoding irrespective of event content.
Figures
Similar articles
-
Medial temporal lobe activity during complex discrimination of faces, objects, and scenes: Effects of viewpoint.Hippocampus. 2010 Mar;20(3):389-401. doi: 10.1002/hipo.20641. Hippocampus. 2010. PMID: 19499575 Free PMC article.
-
Encoding of novel picture pairs activates the perirhinal cortex: an fMRI study.Hippocampus. 2003;13(1):67-80. doi: 10.1002/hipo.10049. Hippocampus. 2003. PMID: 12625459
-
A critical role for the hippocampus and perirhinal cortex in perceptual learning of scenes and faces: complementary findings from amnesia and FMRI.J Neurosci. 2013 Jun 19;33(25):10490-502. doi: 10.1523/JNEUROSCI.2958-12.2013. J Neurosci. 2013. PMID: 23785161 Free PMC article.
-
Visual perception and memory: a new view of medial temporal lobe function in primates and rodents.Annu Rev Neurosci. 2007;30:99-122. doi: 10.1146/annurev.neuro.29.051605.113046. Annu Rev Neurosci. 2007. PMID: 17417938 Review.
-
[Functional neuroimaging studies of episodic memory--functional dissociation in the medial temporal lobe structures].Brain Nerve. 2008 Jul;60(7):833-44. Brain Nerve. 2008. PMID: 18646623 Review. Japanese.
Cited by
-
A Working Memory Buffer in Parahippocampal Regions: Evidence from a Load Effect during the Delay Period.Cereb Cortex. 2016 May;26(5):1965-74. doi: 10.1093/cercor/bhv013. Epub 2015 Feb 6. Cereb Cortex. 2016. PMID: 25662713 Free PMC article.
-
Re-engaging with the past: recapitulation of encoding operations during episodic retrieval.Front Hum Neurosci. 2014 May 27;8:351. doi: 10.3389/fnhum.2014.00351. eCollection 2014. Front Hum Neurosci. 2014. PMID: 24904386 Free PMC article.
-
Complementary attentional components of successful memory encoding.Neuroimage. 2013 Feb 1;66:553-62. doi: 10.1016/j.neuroimage.2012.10.053. Epub 2012 Oct 27. Neuroimage. 2013. PMID: 23108276 Free PMC article.
-
Distinct familiarity-based response patterns for faces and buildings in perirhinal and parahippocampal cortex.J Neurosci. 2013 Jun 26;33(26):10915-23. doi: 10.1523/JNEUROSCI.0126-13.2013. J Neurosci. 2013. PMID: 23804111 Free PMC article.
-
Medial temporal lobe contributions to short-term memory for faces.J Exp Psychol Gen. 2013 Nov;142(4):1309-22. doi: 10.1037/a0033612. Epub 2013 Aug 12. J Exp Psychol Gen. 2013. PMID: 23937185 Free PMC article.
References
-
- Amaral DG, Insausti R. Hippocampal formation. In: Paxinos G, editor. The human nervous system. San Diego, CA: Academic Press; 1990. pp. 711–755.
-
- Amaral DG, Insausti R, Cowan WM. The entorhinal cortex of the monkey: I. Cytoarchitectonic organization. Journal of Comparative Neurology. 1987;264:326–355. - PubMed
-
- Aminoff E, Gronau N, Bar M. The parahippocampal cortex mediates spatial and nonspatial associations. Cerebral Cortex. 2007;17:1493–1503. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous