

Identification of an rsh gene from a Novosphingobium sp. necessary for quorum-sensing signal accumulation
- PMID: 19201802
- PMCID: PMC2668395
- DOI: 10.1128/JB.01692-08
Identification of an rsh gene from a Novosphingobium sp. necessary for quorum-sensing signal accumulation
Abstract
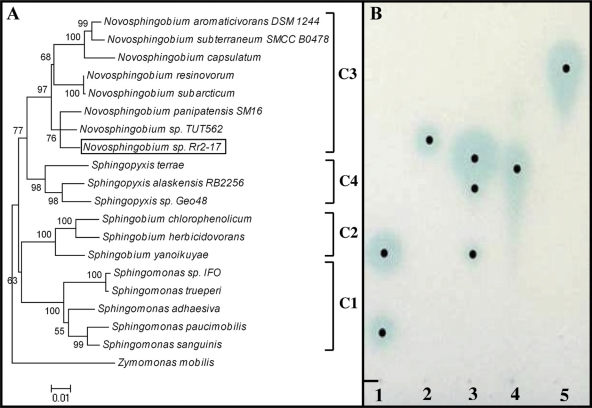
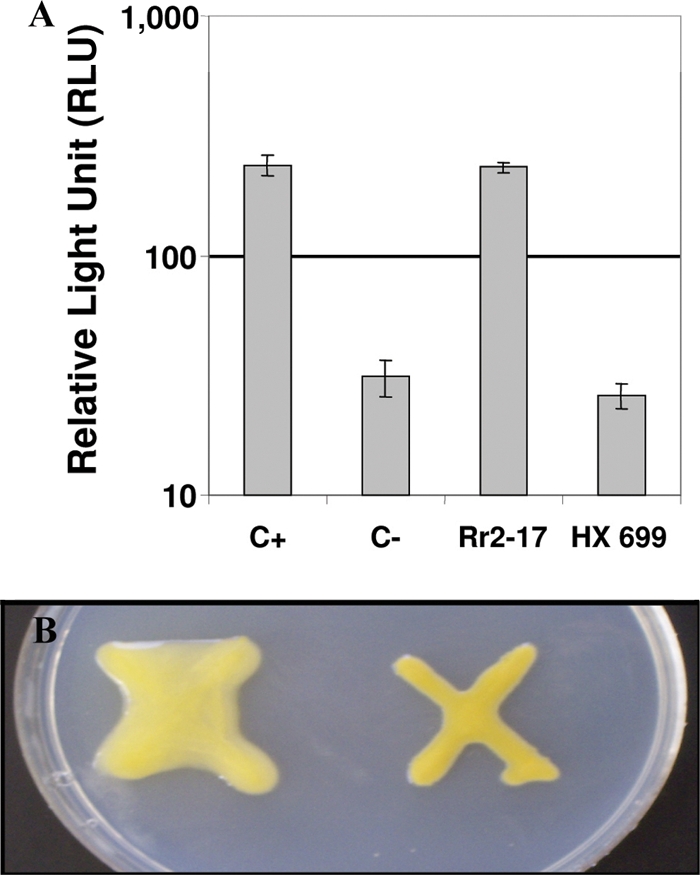
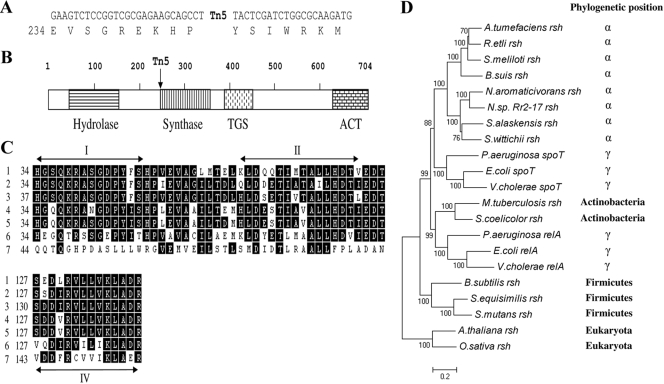
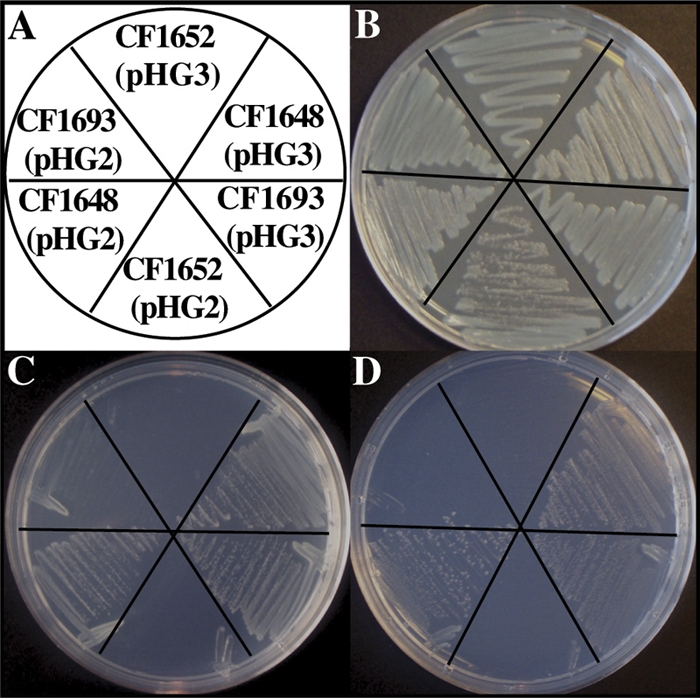
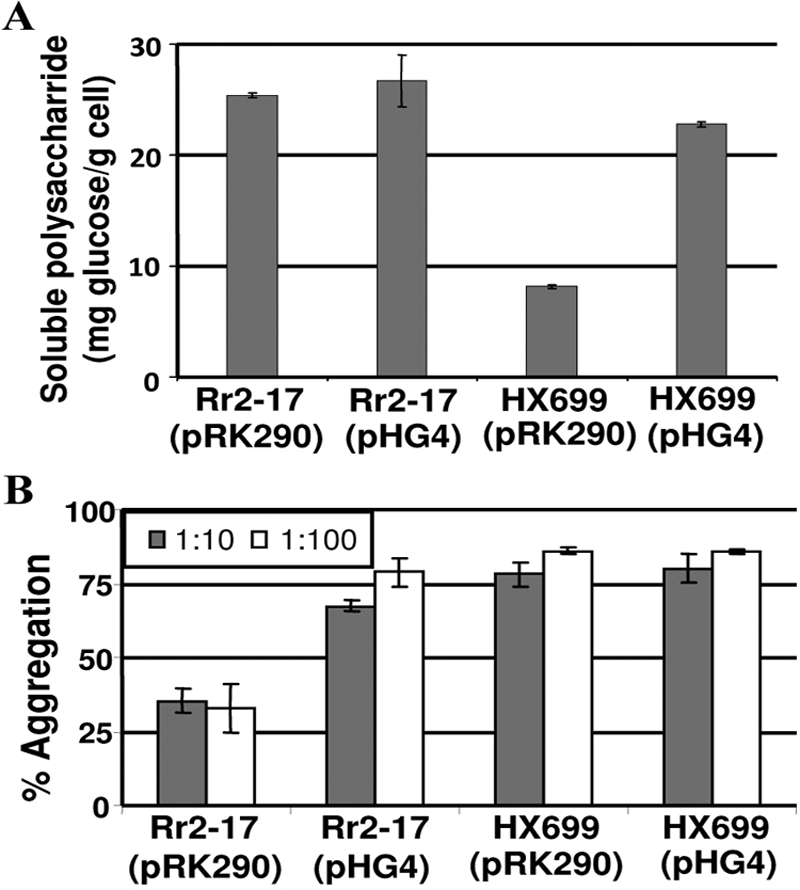
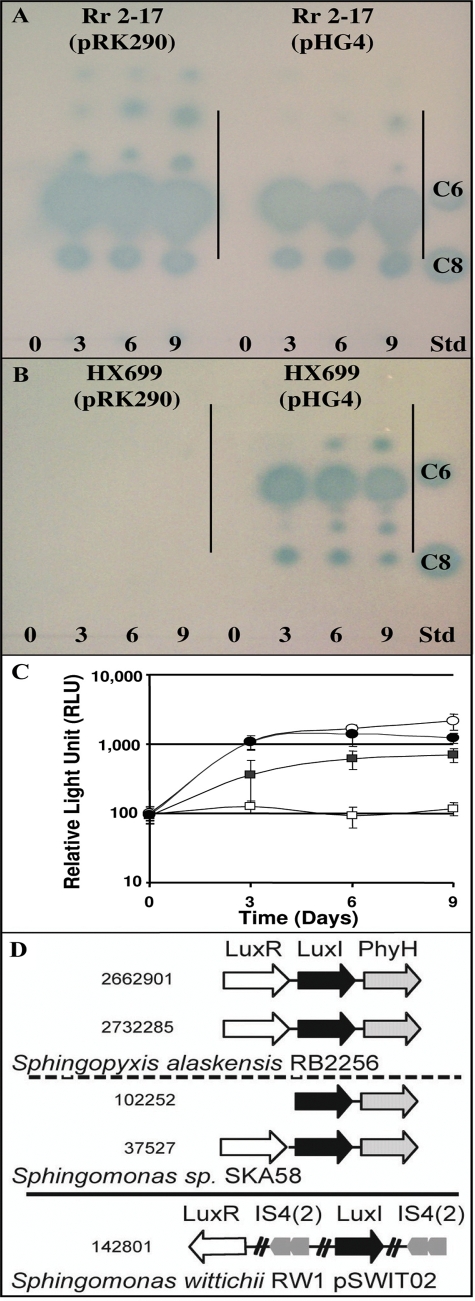
The stringent response is a mechanism by which bacteria adapt to environmental stresses and nutritional deficiencies through the synthesis and hydrolysis of (p)ppGpp by RelA/SpoT enzymes. Alphaproteobacteria and plants contain a single Rsh enzyme (named for RelA/SpoT homolog) that is bifunctional. Here we report the identification of a new species of bacteria belonging to the genus Novosphingobium and characterization of an rsh mutation in this plant tumor-associated isolate. Isolate Rr 2-17, from a grapevine crown gall tumor, is a member of the Novosphingobium genus that produces the N-acyl-homoserine lactone (AHL) quorum-sensing (QS) signals. A Tn5 mutant, Hx 699, deficient in AHL production was found to have an insertion in an rsh gene. The Rsh protein showed significant percent sequence identity to Rsh proteins of alphaproteobacteria. The Novosphingobium sp. rsh gene (rsh(Nsp)) complemented the multiple amino acid requirements of the Escherichia coli relA spoT double mutant by restoring the growth on selection media. Besides QS signal production, the rsh mutation also affects soluble polysaccharide production and cell aggregation. Genetic complementation of the Hx 699 mutant with the rsh(Nsp) gene restored these phenotypes. This is the first discovery of a functional rsh gene in a member of the Novosphingobium genus.
Figures
References
-
- Aravind, L., and E. V. Koonin. 1998. The HD domain defines a new superfamily of metal dependent phosphohydrolases. Trends Biochem. Sci. 23469-472. - PubMed
-
- Braeken, K., M. Moris, R. Daniels, J. Vanderleyden, and J. Michiels. 2006. New horizons for (p)ppGpp in bacterial and plant physiology. Trends Microbiol. 1445-54. - PubMed
-
- Burr, T. J., C. Bazzi, S. Süle, and L. Otten. 1998. Crown gall of grape: biology of Agrobacterium vitis and the development of disease control strategies. Plant Dis. 821288-1297. - PubMed
-
- Calderon-Flores, A., G. Du Pont, A. Huerta-Saquero, H. Merchant-Larios, L. Servin-Gonzalez, and S. Duran. 2005. The stringent response is required for amino acid and nitrate utilization, Nod factor regulation, nodulation, and nitrogen fixation in Rhizobium etli. J. Bacteriol. 1875075-5083. - PMC - PubMed
-
- Case, R. J., M. Labbate, and S. Kejelleberg. 2008. AHL-driven quorum-sensing circuits: their frequency and function among the Proteobacteria. ISME J. 2345-349. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
