

Counting the number of releasable synaptic vesicles in a presynaptic terminal
- PMID: 19202060
- PMCID: PMC2650301
- DOI: 10.1073/pnas.0811017106
Counting the number of releasable synaptic vesicles in a presynaptic terminal
Abstract
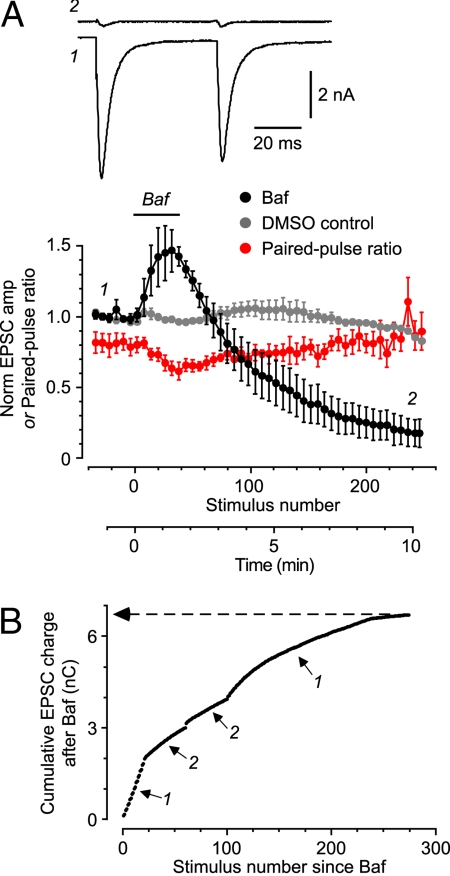
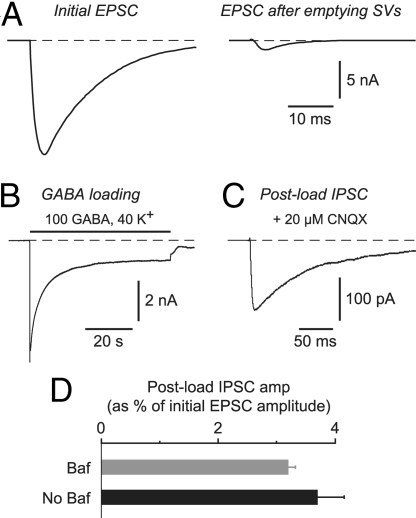
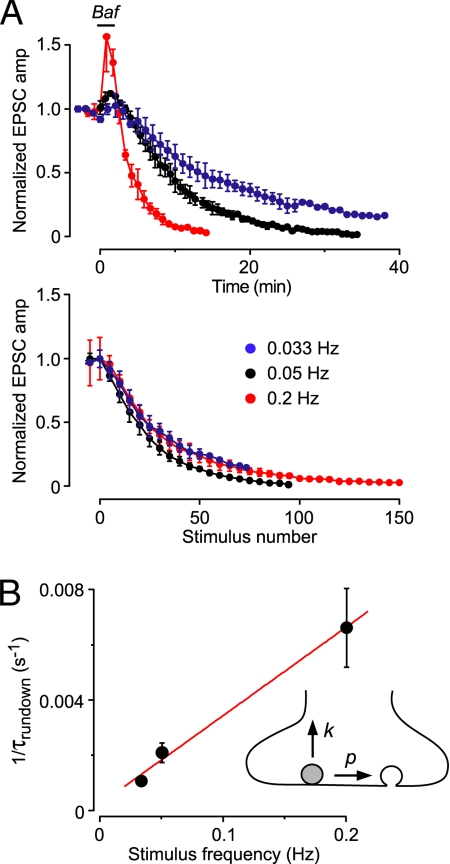
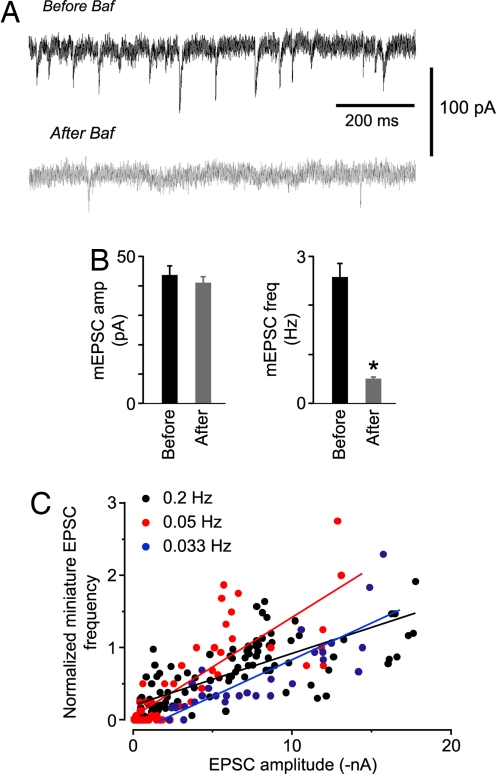
Synaptic transmission depends on the continued availability of neurotransmitter-filled synaptic vesicles (SVs) for triggered release from presynaptic boutons. Surprisingly, small boutons in the brain, that already contain comparatively few SVs, are thought to retain the majority of these SVs in a "reserve" pool that is not mobilized under physiological conditions. Why such a scarce synaptic resource is normally inaccessible has been a matter of debate. Here, we readdress this issue by developing an electrophysiological approach for counting SVs released from boutons formed by a single, isolated neuron on itself ("autapses"). We show that, after treatment with Bafilomycin A1 to prevent reloading of discharged SVs with glutamate, each SV is counted only once on first-time release. Hence, by integrating all autaptic currents as they run down over time, we can estimate the total number of SVs released by a single neuron. This total can be normalized to the number of boutons on the neuron, giving the mean number of SVs released per bouton. We estimate that up to approximately 130 vesicles can be released per bouton over approximately 10 min of stimulation at 0.2 Hz. This number of vesicles represents a substantial proportion of the total number of SVs (100-200) that have been counted in these boutons by using electron microscopy. Thus, mild electrical stimulation, when maintained for sufficient time, causes the eventual release of many of the SVs in a bouton, including those in the putative reserve pool. This result suggests that SVs are functionally homogeneous in that the majority can contribute to basal synaptic transmission.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Rizzoli SO, Betz WJ. Synaptic vesicle pools. Nat Rev Neurosci. 2005;6:57–69. - PubMed
-
- Schweizer FE, Ryan TA. The synaptic vesicle: Cycle of exocytosis and endocytosis. Curr Opin Neurobiol. 2006;16:298–304. - PubMed
-
- Fernandez-Alfonso T, Ryan TA. The efficiency of the synaptic vesicle cycle at central nervous system synapses. Trends Cell Biol. 2006;16:413–420. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
