

Integration of transport-based models for phyllotaxis and midvein formation
- PMID: 19204121
- PMCID: PMC2648550
- DOI: 10.1101/gad.497009
Integration of transport-based models for phyllotaxis and midvein formation
Abstract
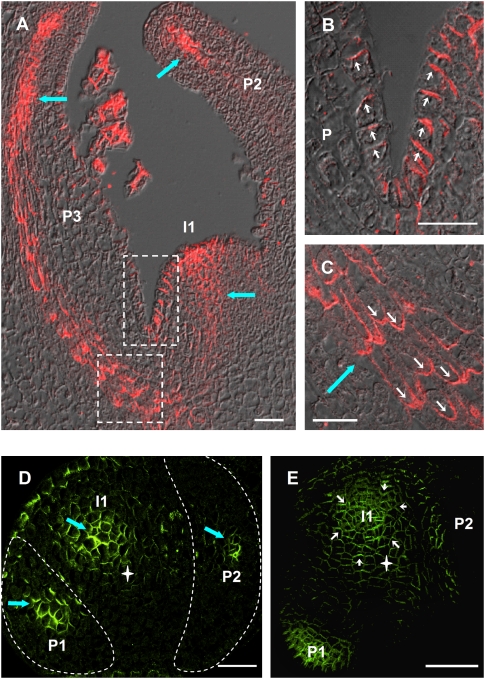
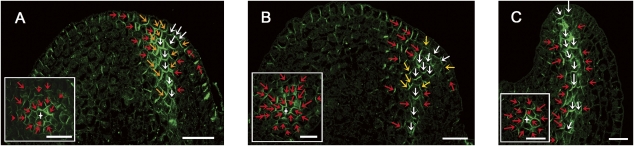
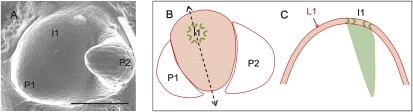
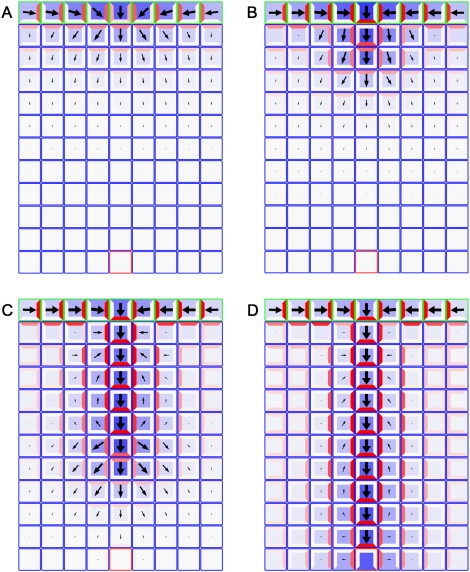
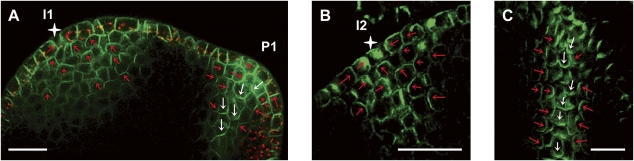
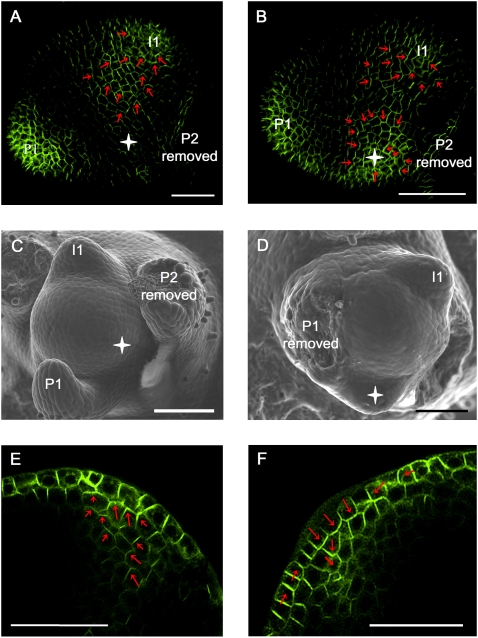
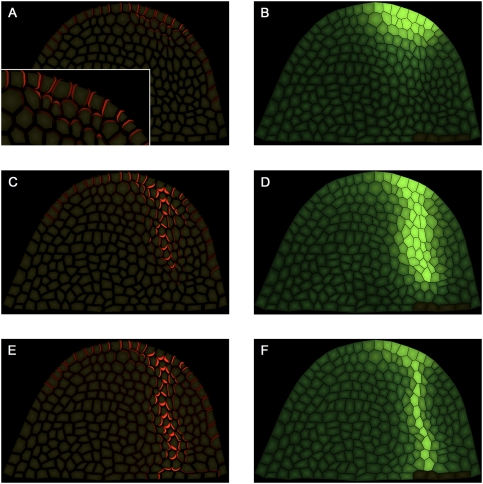
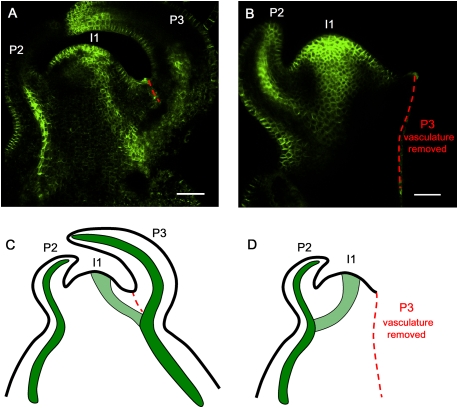
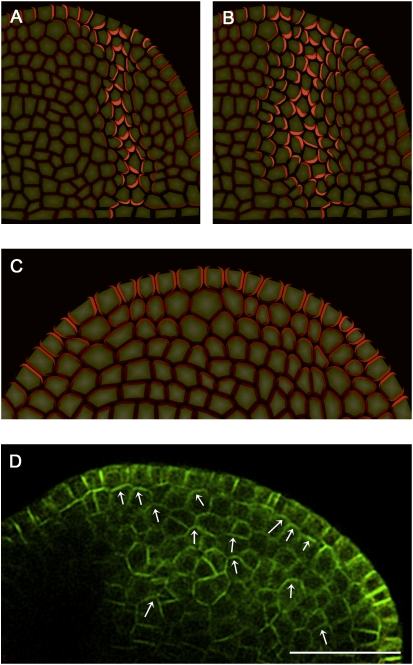
The plant hormone auxin mediates developmental patterning by a mechanism that is based on active transport. In the shoot apical meristem, auxin gradients are thought to be set up through a feedback loop between auxin and the activity and polar localization of its transporter, the PIN1 protein. Two distinct molecular mechanisms for the subcellular polarization of PIN1 have been proposed. For leaf positioning (phyllotaxis), an "up-the-gradient" PIN1 polarization mechanism has been proposed, whereas the formation of vascular strands is thought to proceed by "with-the-flux" PIN1 polarization. These patterning mechanisms intersect during the initiation of the midvein, which raises the question of how two different PIN1 polarization mechanisms may work together. Our detailed analysis of PIN1 polarization during midvein initiation suggests that both mechanisms for PIN1 polarization operate simultaneously. Computer simulations of the resulting dual polarization model are able to reproduce the dynamics of observed PIN1 localization. In addition, the appearance of high auxin concentration in our simulations throughout the initiation of the midvein is consistent with experimental observation and offers an explanation for a long-standing criticism of the canalization hypothesis; namely, how both high flux and high concentration can occur simultaneously in emerging veins.
Figures
References
-
- Baima S., Nobili F., Sessa G., Lucchetti S., Ruberti I., Morelli G. The expression of the Athb-8 homeobox gene is restricted to provascular cells in Arabidopsis thaliana. Development. 1995;121:4171–4182. - PubMed
-
- Benkova E., Michniewicz M., Sauer M., Teichmann T., Seifertova D., Jurgens G., Friml J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell. 2003;115:591–602. - PubMed
-
- Couder Y., Pauchard L., Allain C., Adda-Bedia M., Douady S. The leaf venation as formed in a tensorial field. Eur. Phys. J. B. 2002;28:135–138.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous