

Antiviral immunity in Drosophila requires systemic RNA interference spread
- PMID: 19204732
- PMCID: PMC3978076
- DOI: 10.1038/nature07712
Antiviral immunity in Drosophila requires systemic RNA interference spread
Abstract
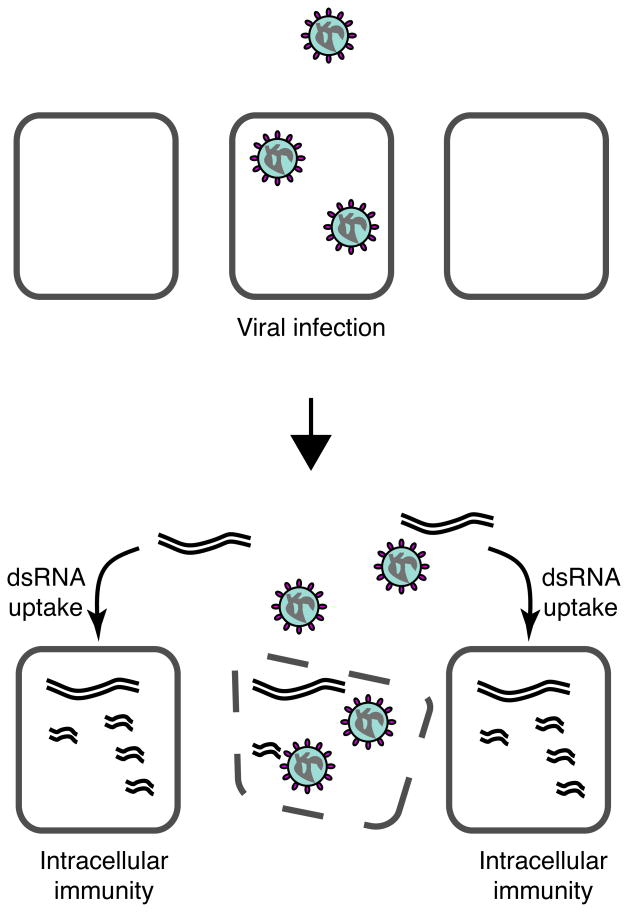
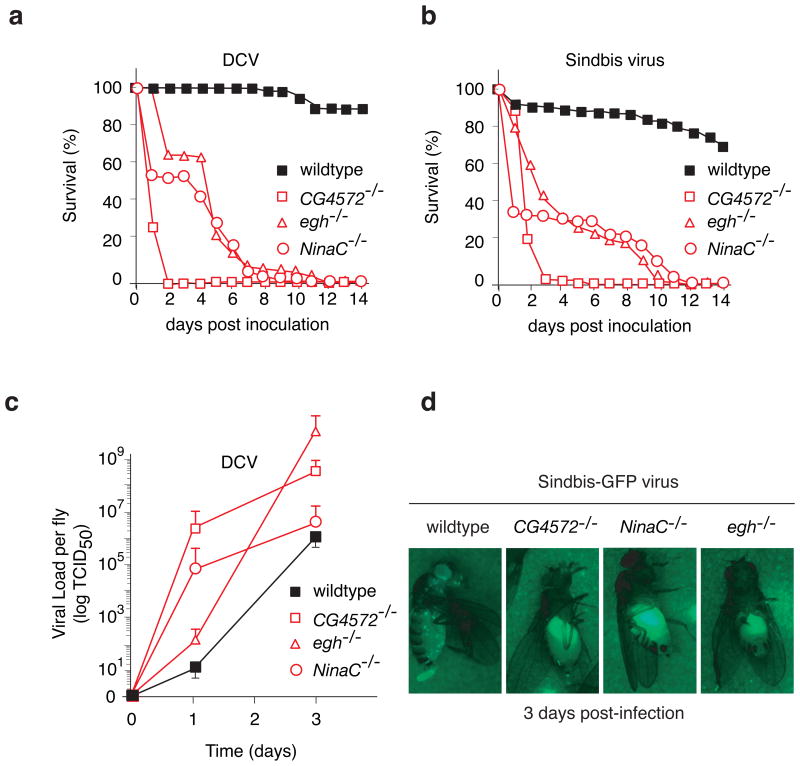
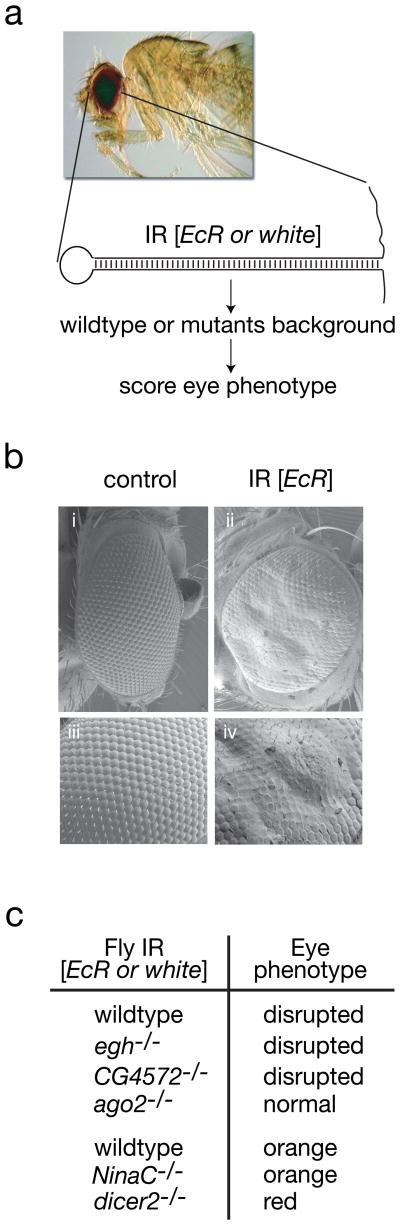
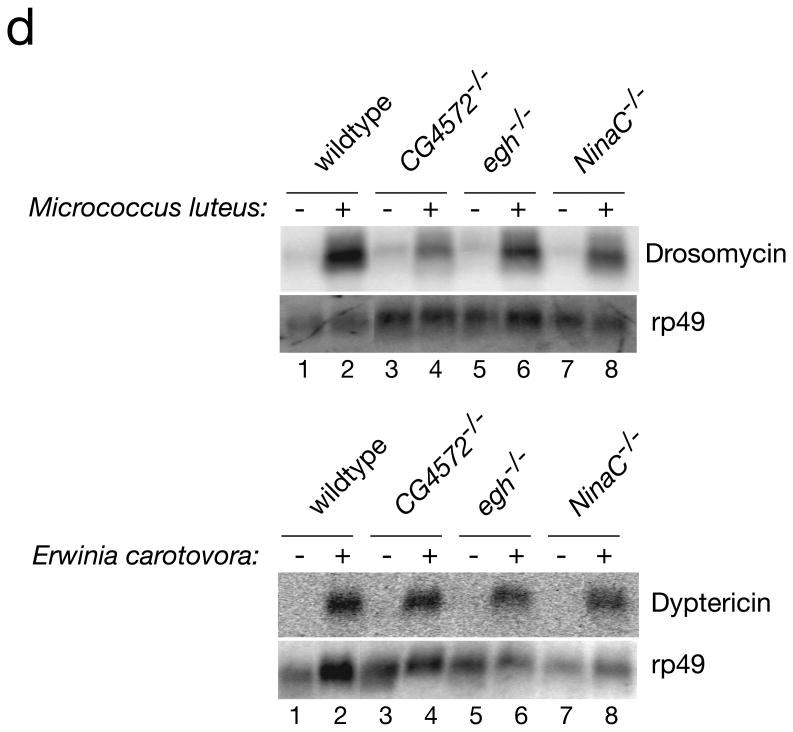
Multicellular organisms evolved sophisticated defence systems to confer protection against pathogens. An important characteristic of these immune systems is their ability to act both locally at the site of infection and at distal uninfected locations. In insects, such as Drosophila melanogaster, RNA interference (RNAi) mediates antiviral immunity. However, the antiviral RNAi defence in flies seems to be a local, cell-autonomous process, as flies are thought to be unable to generate a systemic RNAi response. Here we show that a recently defined double-stranded RNA (dsRNA) uptake pathway is essential for effective antiviral RNAi immunity in adult flies. Mutant flies defective in this dsRNA uptake pathway were hypersensitive to infection with Drosophila C virus and Sindbis virus. Mortality in dsRNA-uptake-defective flies was accompanied by 100-to 10(5)-fold increases in viral titres and higher levels of viral RNA. Furthermore, inoculating naked dsRNA into flies elicited a sequence-specific antiviral immune response that required an intact dsRNA uptake pathway. These findings suggest that spread of dsRNA to uninfected sites is essential for effective antiviral immunity. Notably, infection with green fluorescent protein (GFP)-tagged Sindbis virus suppressed expression of host-encoded GFP at a distal site. Thus, similar to protein-based immunity in vertebrates, the antiviral RNAi response in flies also relies on the systemic spread of a virus-specific immunity signal.
Conflict of interest statement
Figures


References
-
- Baulcombe D. RNA silencing in plants. Nature. 2004;431:356–63. - PubMed
-
- Dorner T, Radbruch A. Antibodies and B cell memory in viral immunity. Immunity. 2007;27:384–92. - PubMed
-
- Roitt I, Brostoff J, Male D. Immunology. Mosby; 2001.
-
- Voinnet O. Non-cell autonomous RNA silencing. FEBS Lett. 2005;579:5858–71. - PubMed
-
- Galiana-Arnoux D, Dostert C, Schneemann A, Hoffmann JA, Imler JL. Essential function in vivo for Dicer-2 in host defense against RNA viruses in drosophila. Nat Immunol. 2006;7:590–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
