

Anastellin, the angiostatic fibronectin peptide, is a selective inhibitor of lysophospholipid signaling
- PMID: 19208746
- PMCID: PMC2658630
- DOI: 10.1158/1541-7786.MCR-08-0195
Anastellin, the angiostatic fibronectin peptide, is a selective inhibitor of lysophospholipid signaling
Abstract
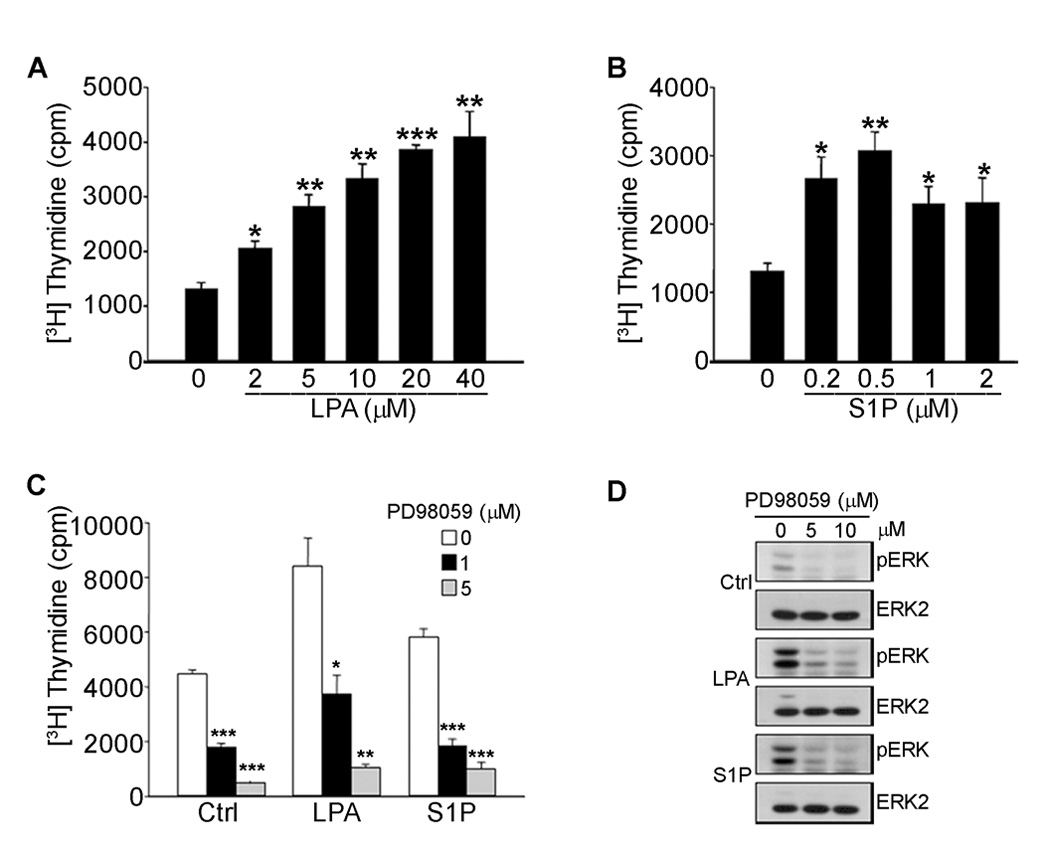
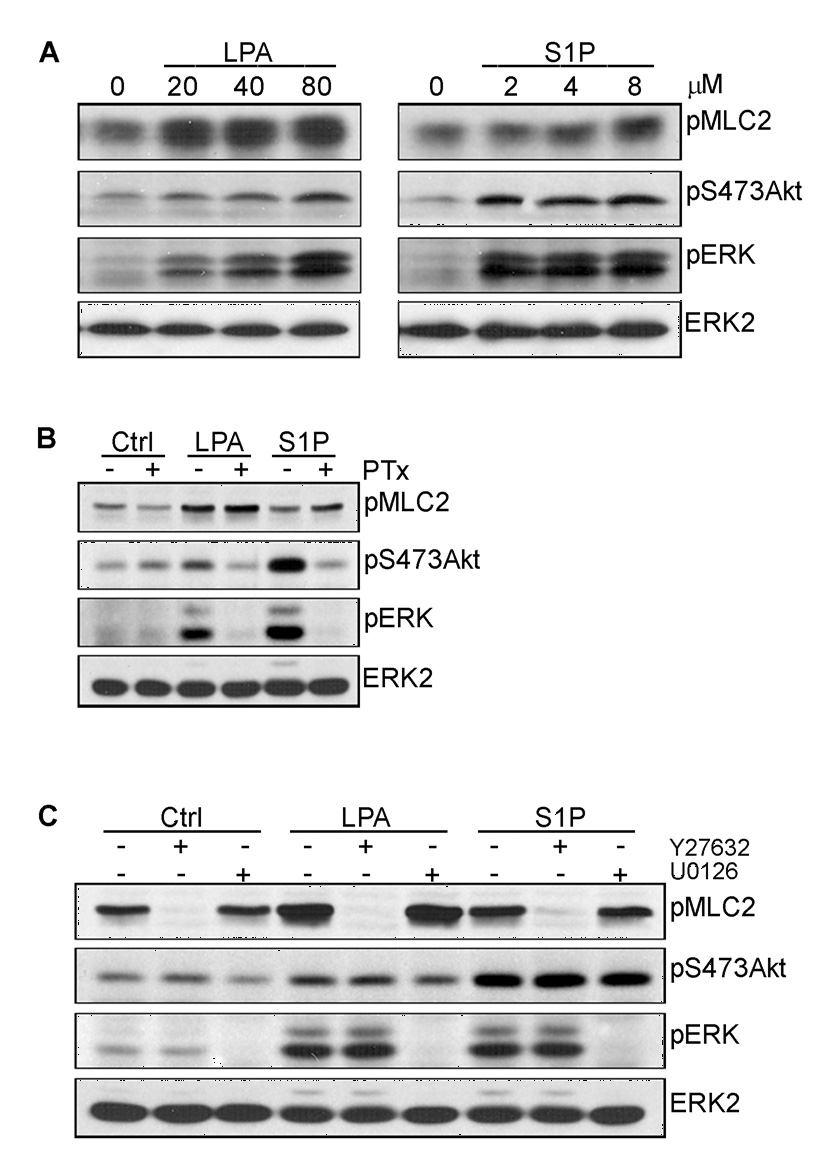
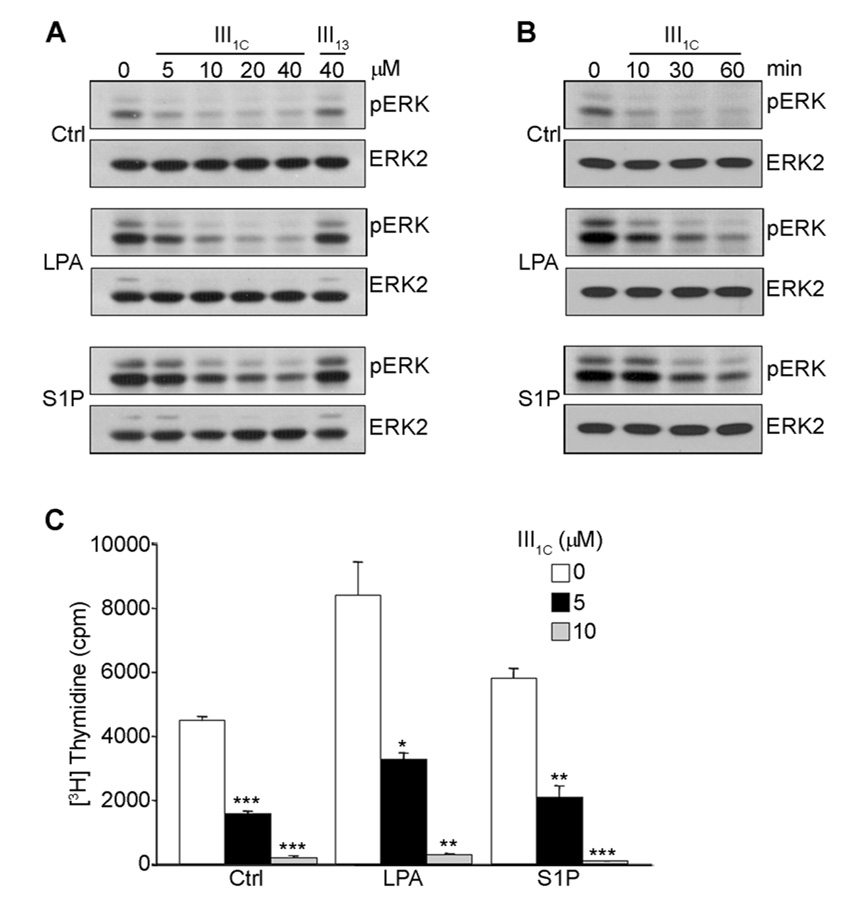
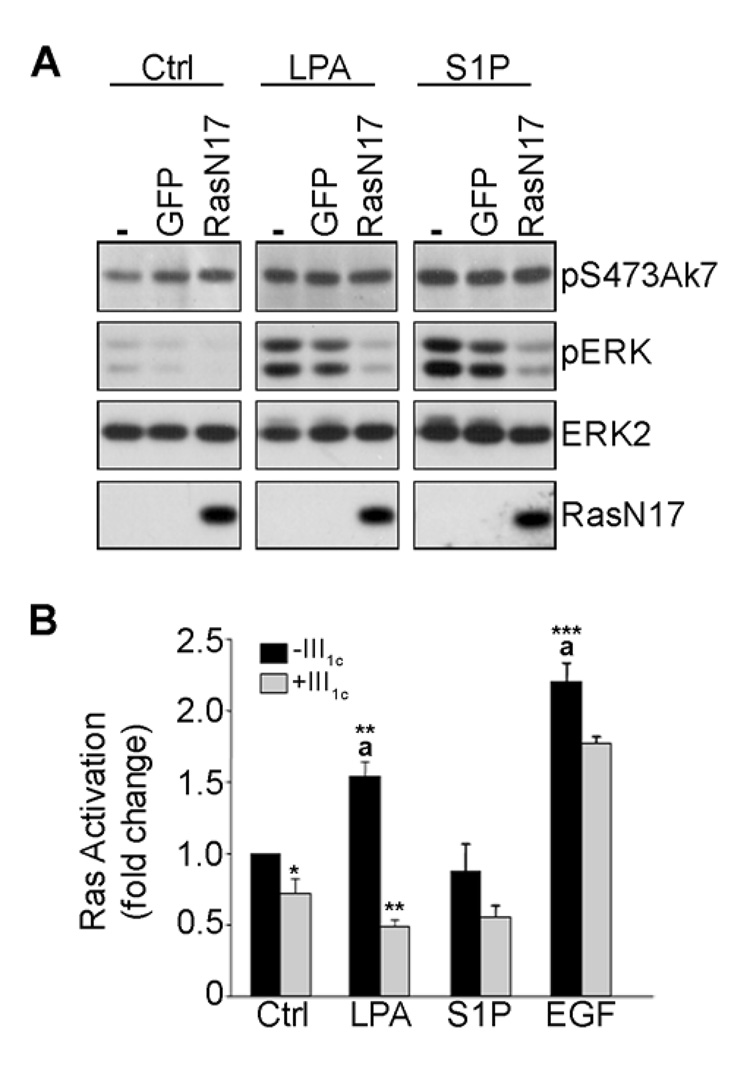
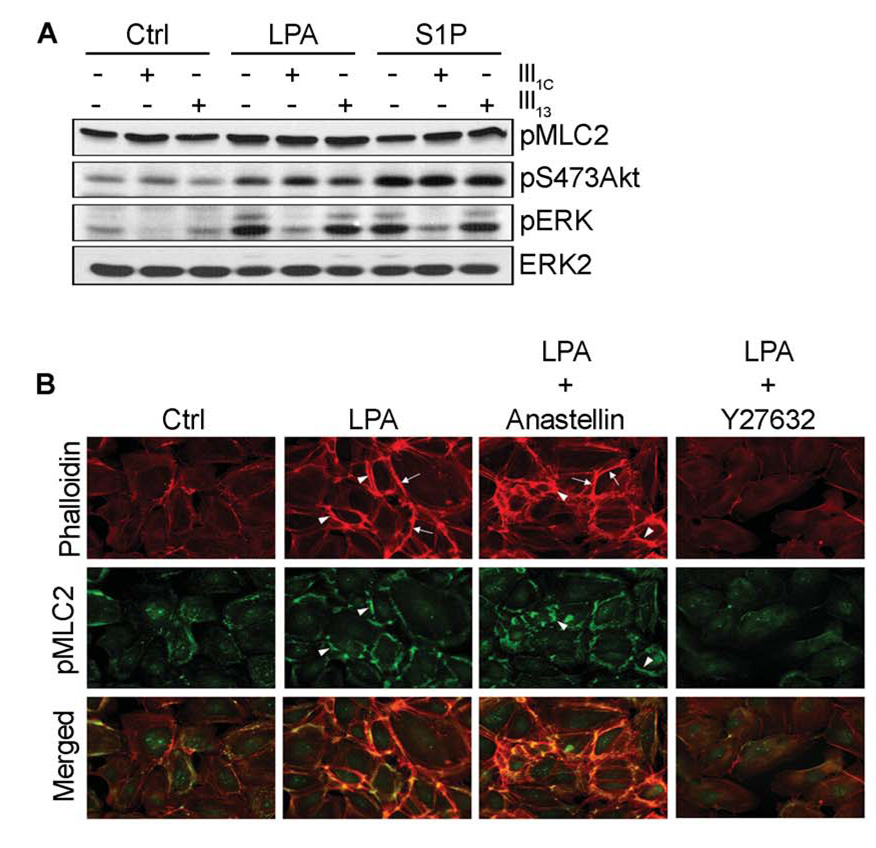
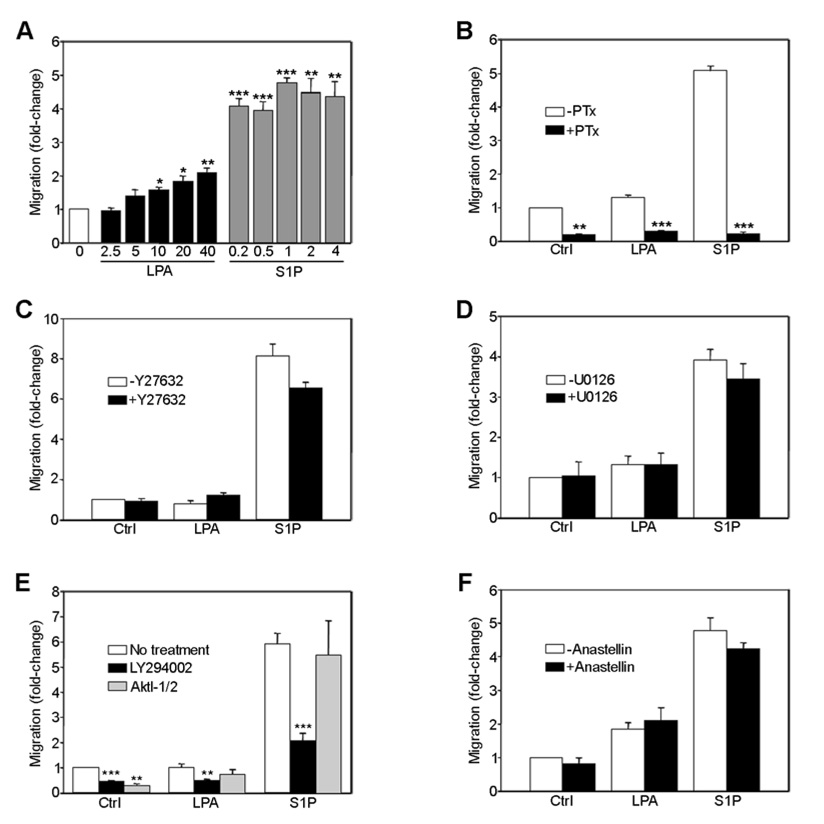
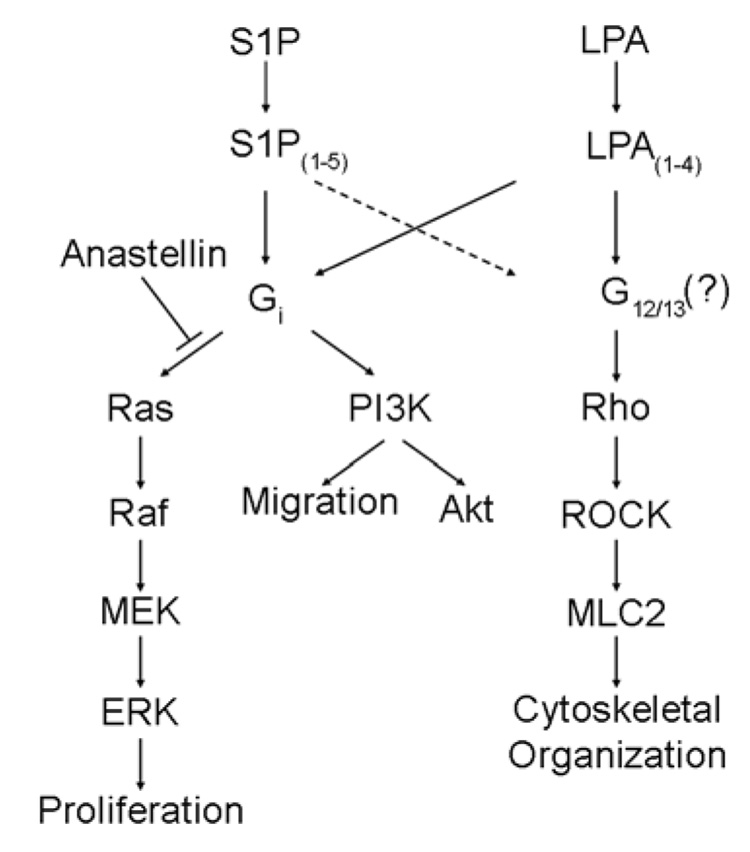
Angiogenesis is regulated by integrin-dependent cell adhesion and the activation of specific cell surface receptors on vascular endothelial cells by angiogenic factors. Lysophosphatidic acid (LPA) and sphingosine-1 phosphate (S1P) are bioactive lysophospholipids that activate G protein-coupled receptors that stimulate phosphatidylinositol 3-kinase (PI3K), Ras, and Rho effector pathways involved in vascular cell survival, proliferation, adhesion, and migration. Previous studies have shown that anastellin, a fragment of the first type III module of fibronectin, functions as an antiangiogenic peptide suppressing tumor growth and metastasis. We have previously shown that anastellin blocks serum-dependent proliferation of microvessel endothelial cells (MVEC) by affecting extracellular signal-regulated kinase (ERK)-dependent G(1)-S transition. However, the mechanism by which anastellin regulates endothelial cell function remains unclear. In the present study, we mapped several lysophospholipid-mediated signaling pathways in MVEC and examined the effects of anastellin on LPA- and S1P-induced MVEC proliferation, migration, and cytoskeletal organization. Both LPA and S1P activated PI3K, Ras/ERK, and Rho/Rho kinase pathways, leading to migration, G(1)-S cell cycle progression, and stress fiber formation, respectively. Stimulation of proliferation by LPA/S1P occurred through a G(i)-dependent Ras/ERK pathway, which was independent of growth factor receptors and PI3K and Rho/Rho kinase signaling. Although LPA and S1P activated both PI3K/Akt and Ras/ERK signaling through G(i), anastellin inhibited only the Ras/ERK pathway. Stress fiber formation in response to LPA was dependent on Rho/Rho kinase but independent of G(i) and unaffected by anastellin. These results suggest that lysophospholipid mediators of G(i) activation leading to PI3K/Akt and Ras/ERK signaling bifurcate downstream of G(i) and that anastellin selectively inhibits the Ras/ERK arm of the pathway.
Conflict of interest statement
No potential conflicts of interest were disclosed.
Figures
Similar articles
-
Anastellin, a fragment of the first type III repeat of fibronectin, inhibits extracellular signal-regulated kinase and causes G(1) arrest in human microvessel endothelial cells.Cancer Res. 2005 Jan 1;65(1):148-56. Cancer Res. 2005. PMID: 15665290
-
Sphingosine-1-phosphate and lysophosphatidic acid stimulate endothelial cell migration.Arterioscler Thromb Vasc Biol. 2000 Apr;20(4):1013-9. doi: 10.1161/01.atv.20.4.1013. Arterioscler Thromb Vasc Biol. 2000. PMID: 10764666
-
Differential regulation of sphingosine-1-phosphate- and VEGF-induced endothelial cell chemotaxis. Involvement of G(ialpha2)-linked Rho kinase activity.Am J Respir Cell Mol Biol. 2001 Jun;24(6):711-9. doi: 10.1165/ajrcmb.24.6.4323. Am J Respir Cell Mol Biol. 2001. PMID: 11415936
-
Sphingosine-1-phosphate signaling in physiology and diseases.Biofactors. 2012 Sep-Oct;38(5):329-37. doi: 10.1002/biof.1030. Epub 2012 Jun 7. Biofactors. 2012. PMID: 22674845 Review.
-
G protein mediated signaling pathways in lysophospholipid induced cell proliferation and survival.J Cell Biochem. 2004 Aug 1;92(5):949-66. doi: 10.1002/jcb.20094. J Cell Biochem. 2004. PMID: 15258918 Review.
Cited by
-
Cytosolic phospholipase A2 and lysophospholipids in tumor angiogenesis.J Natl Cancer Inst. 2010 Sep 22;102(18):1398-412. doi: 10.1093/jnci/djq290. Epub 2010 Aug 20. J Natl Cancer Inst. 2010. PMID: 20729478 Free PMC article.
-
Tumor Microenvironment: Extracellular Matrix Alterations Influence Tumor Progression.Front Oncol. 2020 Apr 15;10:397. doi: 10.3389/fonc.2020.00397. eCollection 2020. Front Oncol. 2020. PMID: 32351878 Free PMC article. Review.
-
The First Type III Domain of Fibronectin is Associated with the Expression of Cytokines within the Lung Tumor Microenvironment.J Cancer. 2011;2:478-83. doi: 10.7150/jca.2.478. Epub 2011 Sep 27. J Cancer. 2011. PMID: 21980322 Free PMC article.
-
The first type III repeat in fibronectin activates an inflammatory pathway in dermal fibroblasts.J Biol Chem. 2010 Nov 19;285(47):36255-9. doi: 10.1074/jbc.C110.176990. Epub 2010 Oct 5. J Biol Chem. 2010. PMID: 20923762 Free PMC article.
-
Extracellular matrix dynamics in vascular remodeling.Am J Physiol Cell Physiol. 2020 Sep 1;319(3):C481-C499. doi: 10.1152/ajpcell.00147.2020. Epub 2020 Jun 24. Am J Physiol Cell Physiol. 2020. PMID: 32579472 Free PMC article. Review.
References
-
- Ambesi A, Klein RM, Pumiglia KM, McKeown-Longo PJ. Anastellin, a fragment of the first type III repeat of fibronectin, inhibits extracellular signal-regulated kinase and causes G1 arrest in human microvessel endothelial cells. Cancer Res. 2005;65:148–156. - PubMed
-
- Hynes RO. Cell matrix adhesion in vascular development. J Thromb Haemost. 2007;5 Suppl 1:32–40. - PubMed
-
- Goetzl EJ, An S. Diversity of cellular receptors and functions for the lysophospholipid growth factors lysophosphatidic acid and sphingosine 1-phosphate. FASEB J. 1998;12:1589–1598. - PubMed
-
- Xu Y, Shen Z, Wiper DW, Wu M, Morton RE, Elson P, Kennedy AW, Belinson J, Markman M, Casey G. Lysophosphatidic acid as a potential biomarker for ovarian and other gynecologic cancers. J Am Med Assoc. 1998;280:719–723. - PubMed
-
- Hart JJ, Jiang X, Kozasa T, Roscoe W, Singer WD, Gilman AG, Sternweis PC, Bollag G. Direct stimulation of the guanine nucleotide exchange activity of p115 RhoGEF by Gα13. Science. 1998;280:2112–2114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
