

How (and why) the immune system makes us sleep
- PMID: 19209176
- PMCID: PMC2839418
- DOI: 10.1038/nrn2576
How (and why) the immune system makes us sleep
Abstract
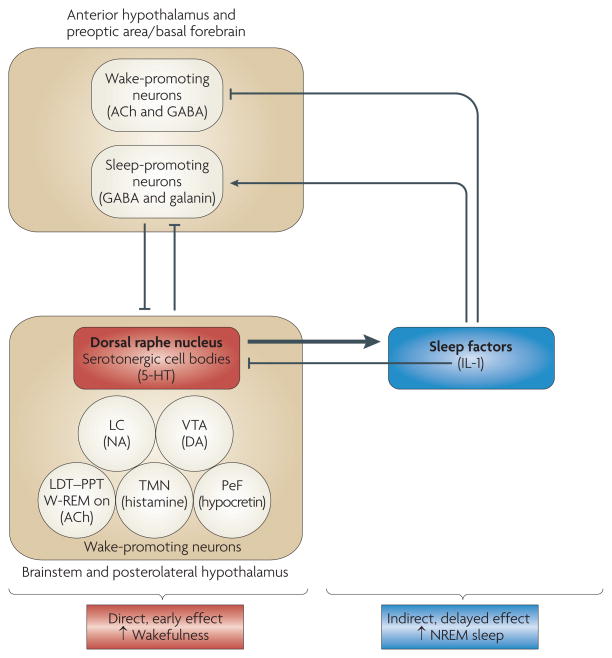
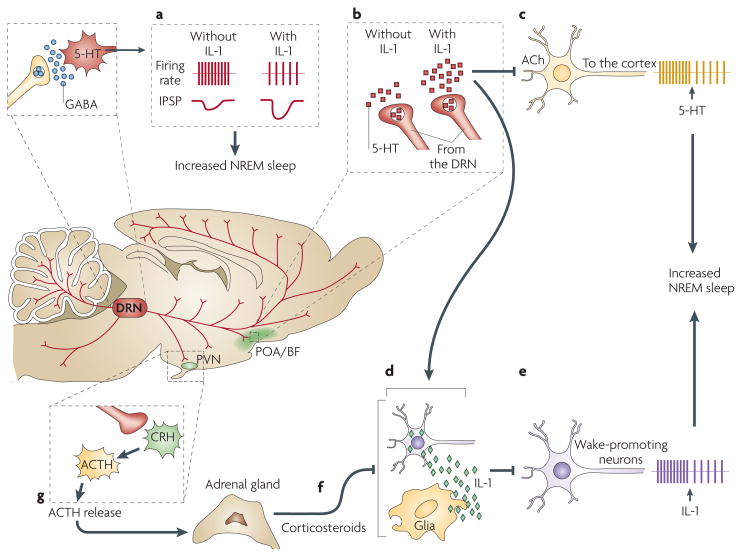
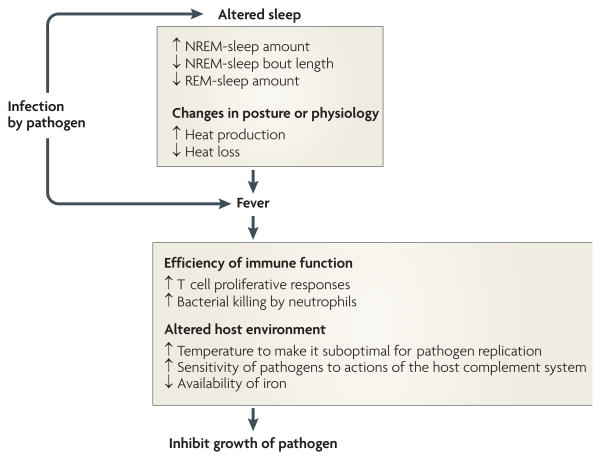
Good sleep is necessary for physical and mental health. For example, sleep loss impairs immune function, and sleep is altered during infection. Immune signalling molecules are present in the healthy brain, where they interact with neurochemical systems to contribute to the regulation of normal sleep. Animal studies have shown that interactions between immune signalling molecules (such as the cytokine interleukin 1) and brain neurochemical systems (such as the serotonin system) are amplified during infection, indicating that these interactions might underlie the changes in sleep that occur during infection. Why should the immune system cause us to sleep differently when we are sick? We propose that the alterations in sleep architecture during infection are exquisitely tailored to support the generation of fever, which in turn imparts survival value.
Figures


References
-
- Tononi G, Cirelli C. Sleep and synaptic homeostasis: a hypothesis. Brain Res Bull. 2003;62:143–150. - PubMed
-
- Centers for Disease Control and Prevention. Percentage of adults who reported an average of ≤6 hours of sleep per 24-hour period, by sex and age group—United States 1985 and 2004. MMWR Morb Mortal Wkly Rep. 2005;54:933.
-
- Spiegel K, Sheridan JF, Van Cauter E. Effect of sleep deprivation on response to immunization. JAMA. 2002;288:1471–1472. - PubMed
